

The R3-Type MYB Transcription Factor BrMYBL2.1 Negatively Regulates Anthocyanin Biosynthesis in Chinese Cabbage (Brassica rapa L.) by Repressing MYB-bHLH-WD40 Complex Activity
- PMID: 35328800
- PMCID: PMC8949199
- DOI: 10.3390/ijms23063382
The R3-Type MYB Transcription Factor BrMYBL2.1 Negatively Regulates Anthocyanin Biosynthesis in Chinese Cabbage (Brassica rapa L.) by Repressing MYB-bHLH-WD40 Complex Activity
Abstract
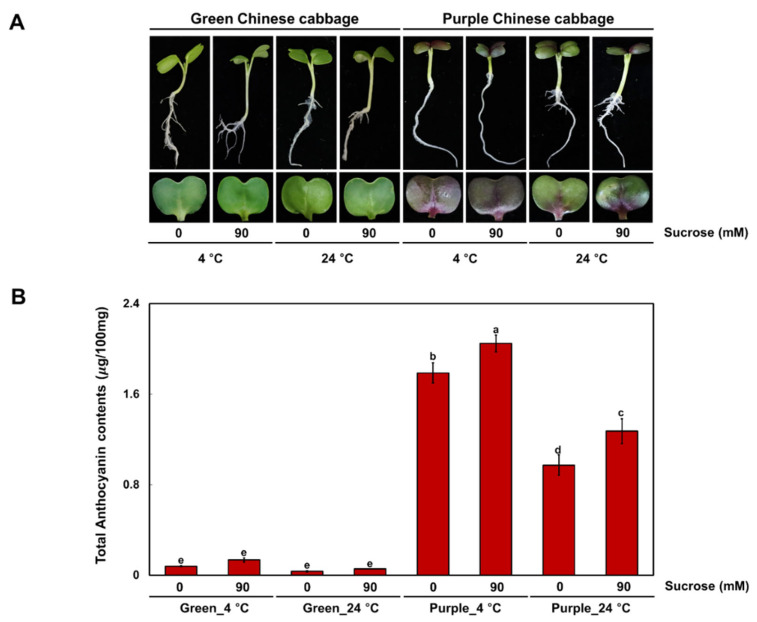
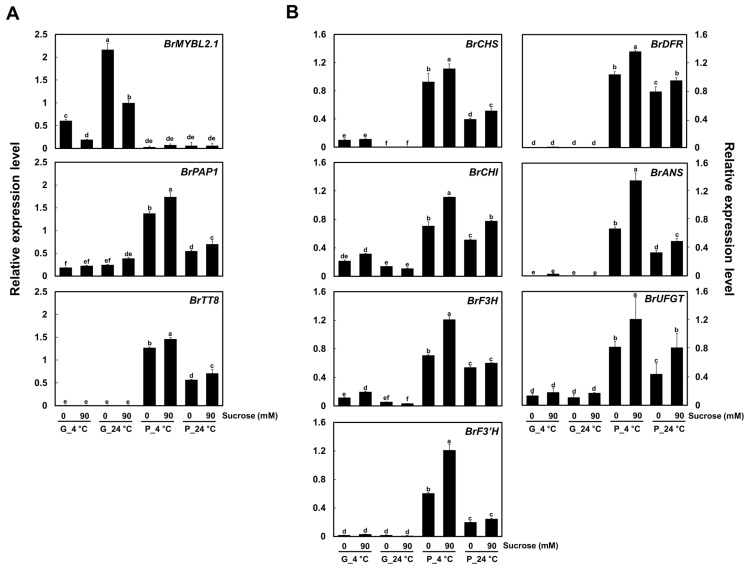
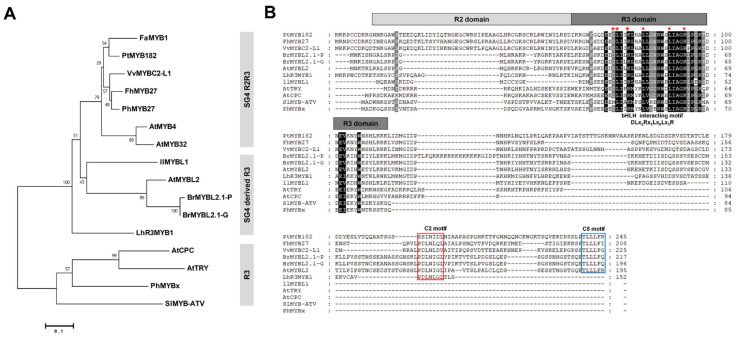
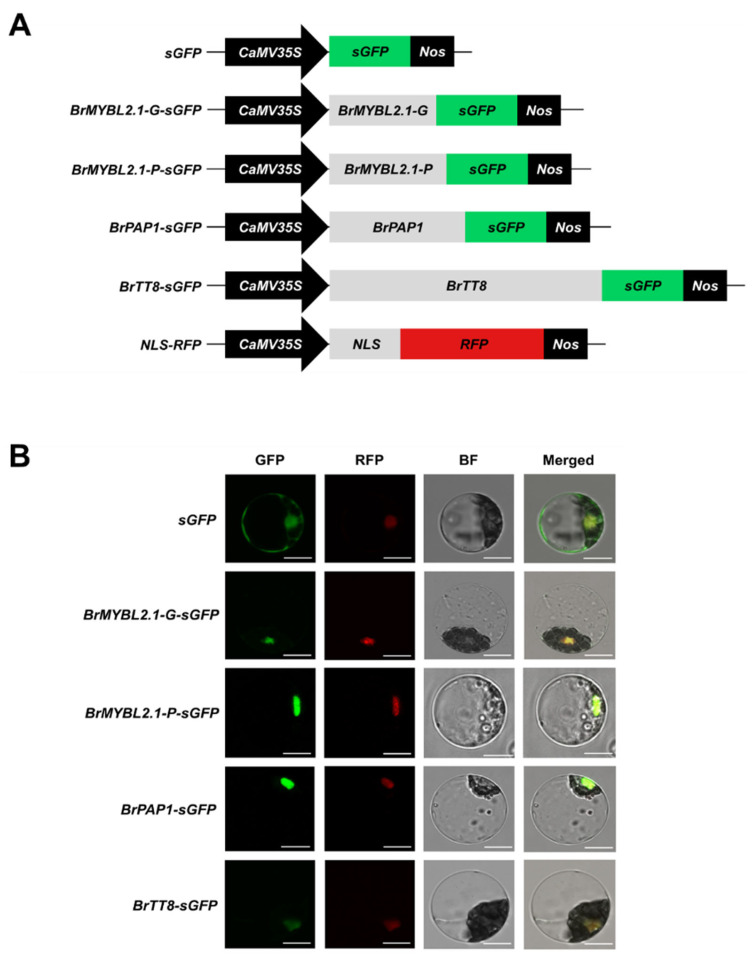
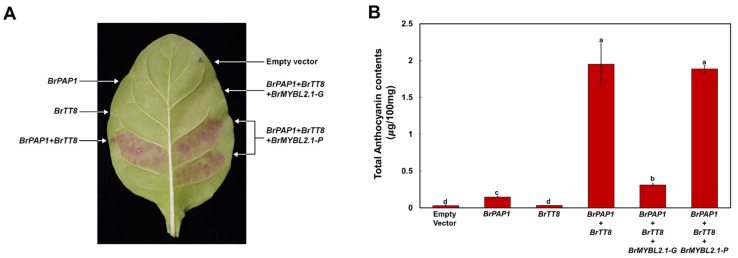
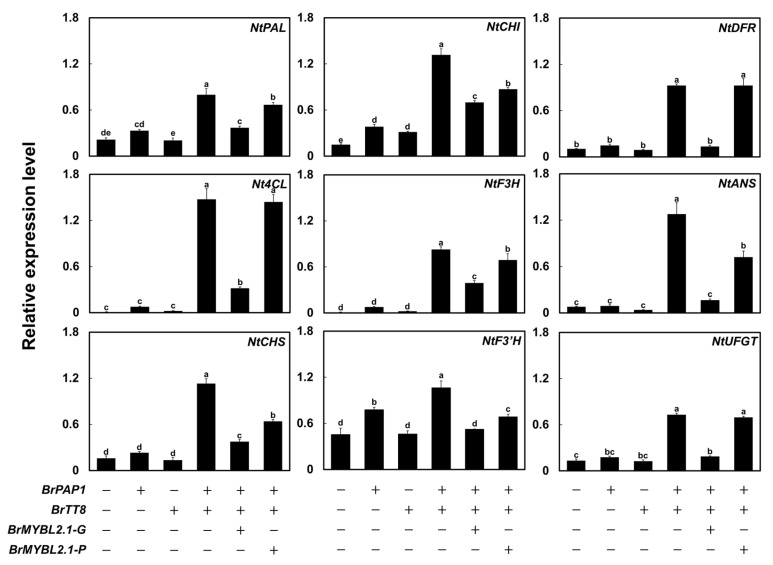
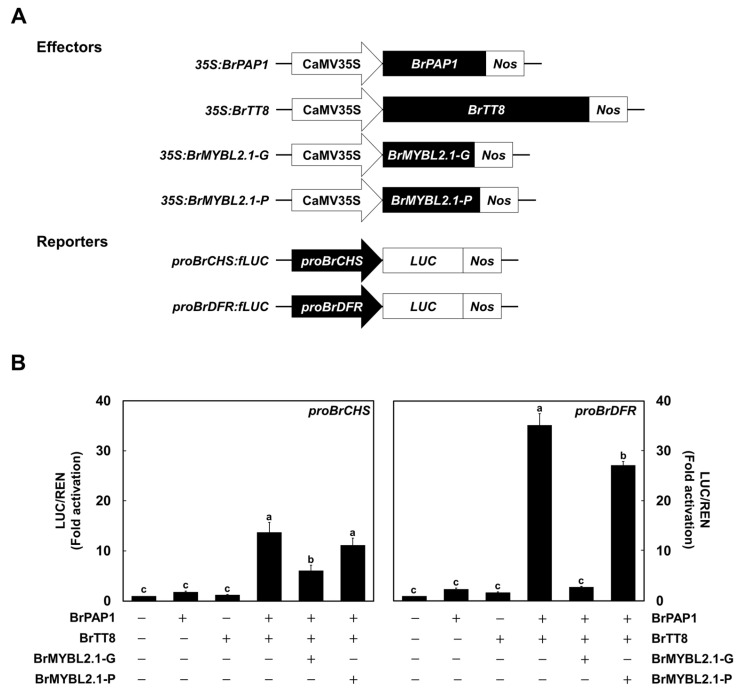
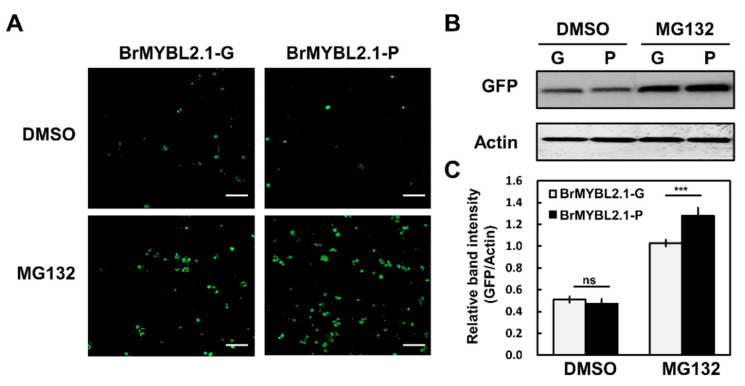
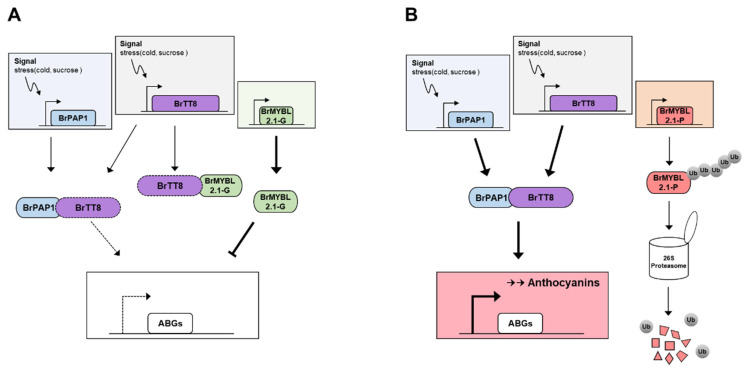
Chinese cabbage (Brassica rapa L.) leaves are purple in color due to anthocyanin accumulation and have nutritional and aesthetic value, as well as antioxidant properties. Here, we identified the R3 MYB transcription factor BrMYBL2.1 as a key negative regulator of anthocyanin biosynthesis. A Chinese cabbage cultivar with green leaves harbored a functional BrMYBL2.1 protein, designated BrMYBL2.1-G, with transcriptional repressor activity of anthocyanin biosynthetic genes. By contrast, BrMYBL2.1 from a Chinese cabbage cultivar with purple leaves carried a poly(A) insertion in the third exon of the gene, resulting in the insertion of multiple lysine residues in the predicted protein, designated BrMYBL2.1-P. Although both BrMYBL2.1 variants localized to the nucleus, only BrMYBL2.1-G interacted with its cognate partner BrTT8. Transient infiltration assays in tobacco leaves revealed that BrMYBL2.1-G, but not BrMYBL2.1-P, actively represses pigment accumulation by inhibiting the transcription of anthocyanin biosynthetic genes. Transient promoter activation assay in Arabidopsis protoplasts verified that BrMYBL2.1-G, but not BrMYBL2.1-P, can repress transcriptional activation of BrCHS and BrDFR, which was activated by co-expression with BrPAP1 and BrTT8. We determined that BrMYBL2.1-P may be more prone to degradation than BrMYBL2.1-G via ubiquitination. Taken together, these results demonstrate that BrMYBL2.1-G blocks the activity of the MBW complex and thus represses anthocyanin biosynthesis, whereas the variant BrMYBL2.1-P from purple Chinese cabbage cannot, thus leading to higher anthocyanin accumulation.
Keywords: BrMYBL2.1; Chinese cabbage; anthocyanin; insertional mutation; repressor; ubiquitination.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
R2R3-MYB repressor, BrMYB32, regulates anthocyanin biosynthesis in Chinese cabbage.Physiol Plant. 2024 Nov-Dec;176(6):e14591. doi: 10.1111/ppl.14591. Physiol Plant. 2024. PMID: 39468991
-
Low Temperature Promotes Anthocyanin Biosynthesis and Related Gene Expression in the Seedlings of Purple Head Chinese Cabbage (Brassica rapa L.).Genes (Basel). 2020 Jan 10;11(1):81. doi: 10.3390/genes11010081. Genes (Basel). 2020. PMID: 31936856 Free PMC article.
-
A single amino acid substitution in the R2R3 conserved domain of the BrPAP1a transcription factor impairs anthocyanin production in turnip (Brassica rapa subsp. rapa).Plant Physiol Biochem. 2021 May;162:124-136. doi: 10.1016/j.plaphy.2021.02.011. Epub 2021 Feb 22. Plant Physiol Biochem. 2021. PMID: 33676299
-
GLABRA2, A Common Regulator for Epidermal Cell Fate Determination and Anthocyanin Biosynthesis in Arabidopsis.Int J Mol Sci. 2019 Oct 9;20(20):4997. doi: 10.3390/ijms20204997. Int J Mol Sci. 2019. PMID: 31601032 Free PMC article. Review.
-
Advance of the negative regulation of anthocyanin biosynthesis by MYB transcription factors.Plant Physiol Biochem. 2019 Mar;136:178-187. doi: 10.1016/j.plaphy.2019.01.024. Epub 2019 Jan 22. Plant Physiol Biochem. 2019. PMID: 30685697 Review.
Cited by
-
Identification and characteristics of differentially expressed genes under UV-B stress in Gossypium hirsutum.Front Plant Sci. 2025 Jan 15;15:1529912. doi: 10.3389/fpls.2024.1529912. eCollection 2024. Front Plant Sci. 2025. PMID: 39881734 Free PMC article.
-
Genome-wide identification of MYBL2 in Brassicaceae, with a focus on the expression pattern of regulating anthocyanin synthesis in Brassica crops.Front Plant Sci. 2025 Jul 1;16:1629560. doi: 10.3389/fpls.2025.1629560. eCollection 2025. Front Plant Sci. 2025. PMID: 40666295 Free PMC article.
-
A Mutation in the MYBL2-1 Gene Is Associated with Purple Pigmentation in Brassica oleracea.Int J Mol Sci. 2022 Oct 6;23(19):11865. doi: 10.3390/ijms231911865. Int J Mol Sci. 2022. PMID: 36233166 Free PMC article.
-
Transcriptome and Metabolome Profiling to Explore the Causes of Purple Leaves Formation in Non-Heading Chinese Cabbage (Brassica rapa L. ssp. chinensis Makino var. mutliceps Hort.).Foods. 2022 Jun 17;11(12):1787. doi: 10.3390/foods11121787. Foods. 2022. PMID: 35741985 Free PMC article.
-
The introgression of BjMYB113 from Brassica juncea leads to purple leaf trait in Brassica napus.BMC Plant Biol. 2024 Aug 2;24(1):735. doi: 10.1186/s12870-024-05418-5. BMC Plant Biol. 2024. PMID: 39090544 Free PMC article.
References
-
- Nakabayashi R., Yonekura-Sakakibara K., Urano K., Suzuki M., Yamada Y., Nishizawa T., Matsuda F., Kojima M., Sakakibara H., Shinozaki K., et al. Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 2014;77:367–379. doi: 10.1111/tpj.12388. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
