

Transcriptional profiling of β-2M-SPα-6+THY1+ spermatogonial stem cells in human spermatogenesis
- PMID: 35334216
- PMCID: PMC9023810
- DOI: 10.1016/j.stemcr.2022.02.017
Transcriptional profiling of β-2M-SPα-6+THY1+ spermatogonial stem cells in human spermatogenesis
Abstract
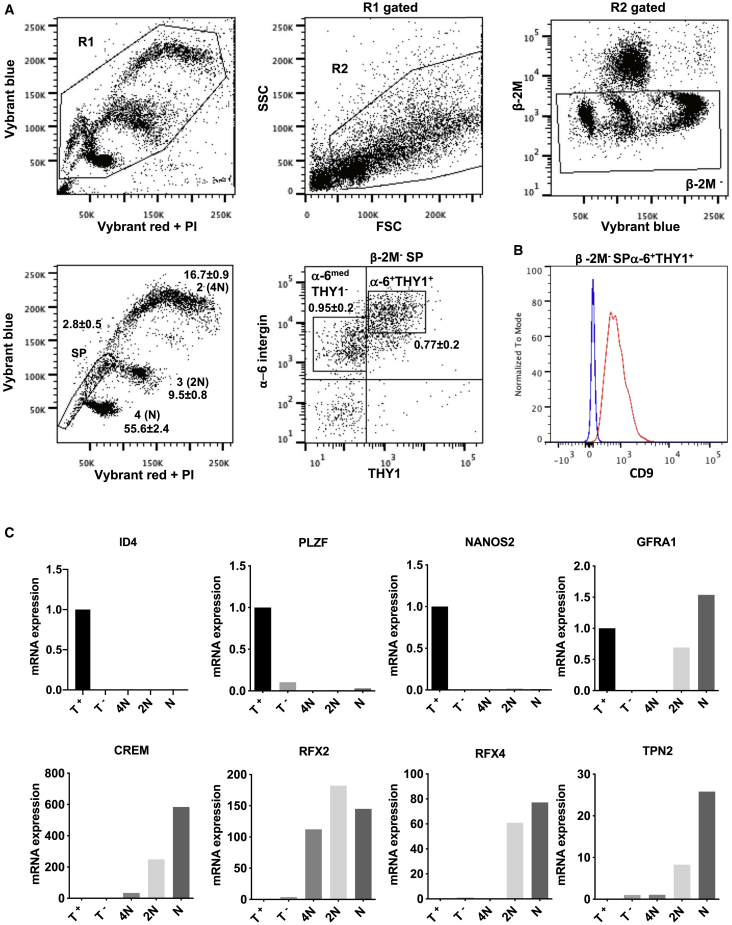
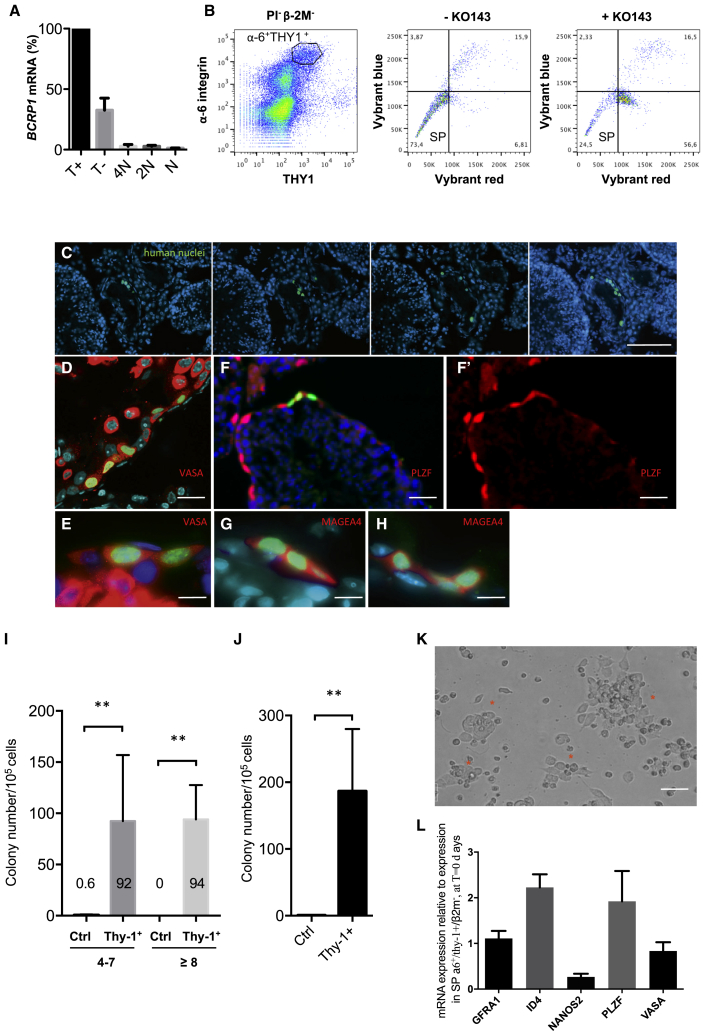
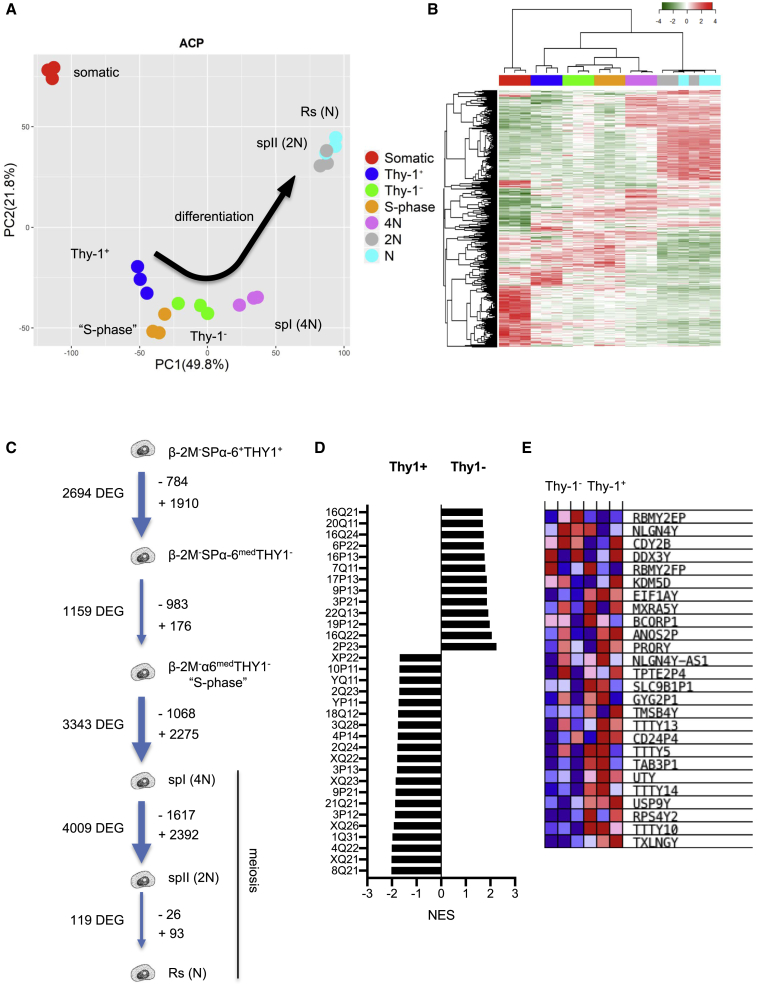
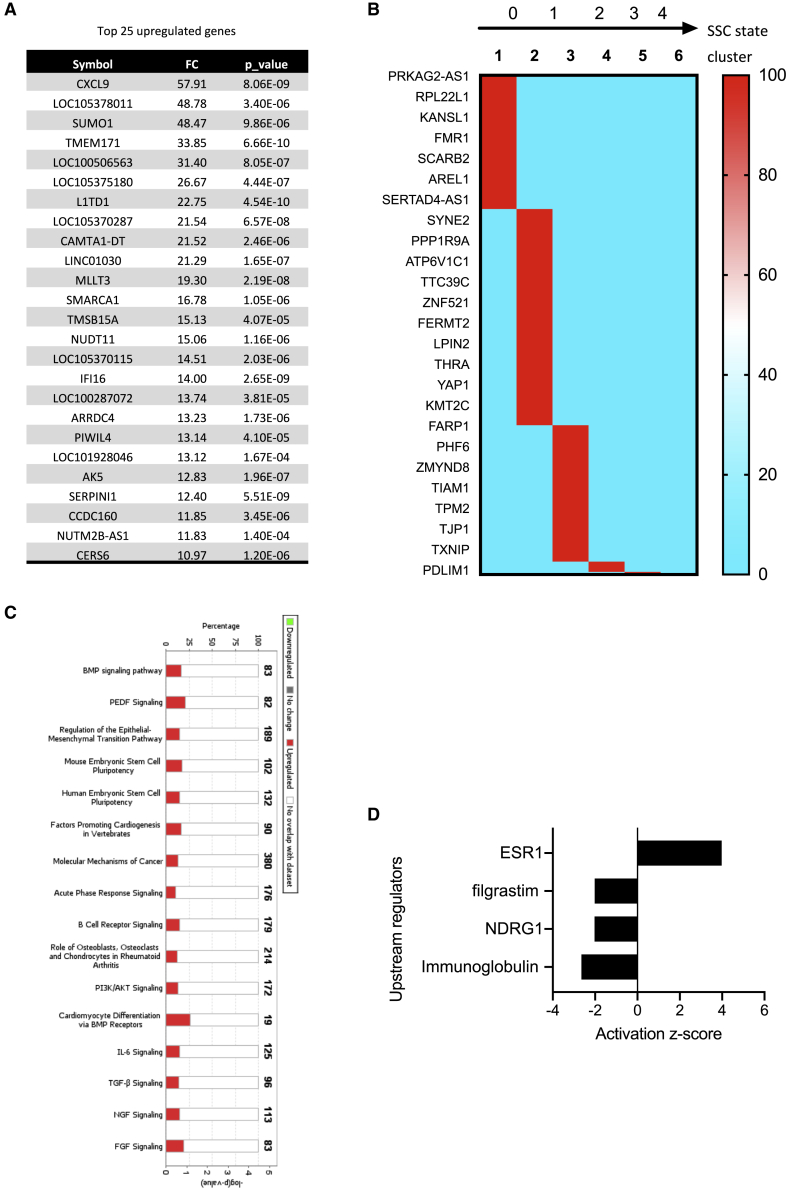
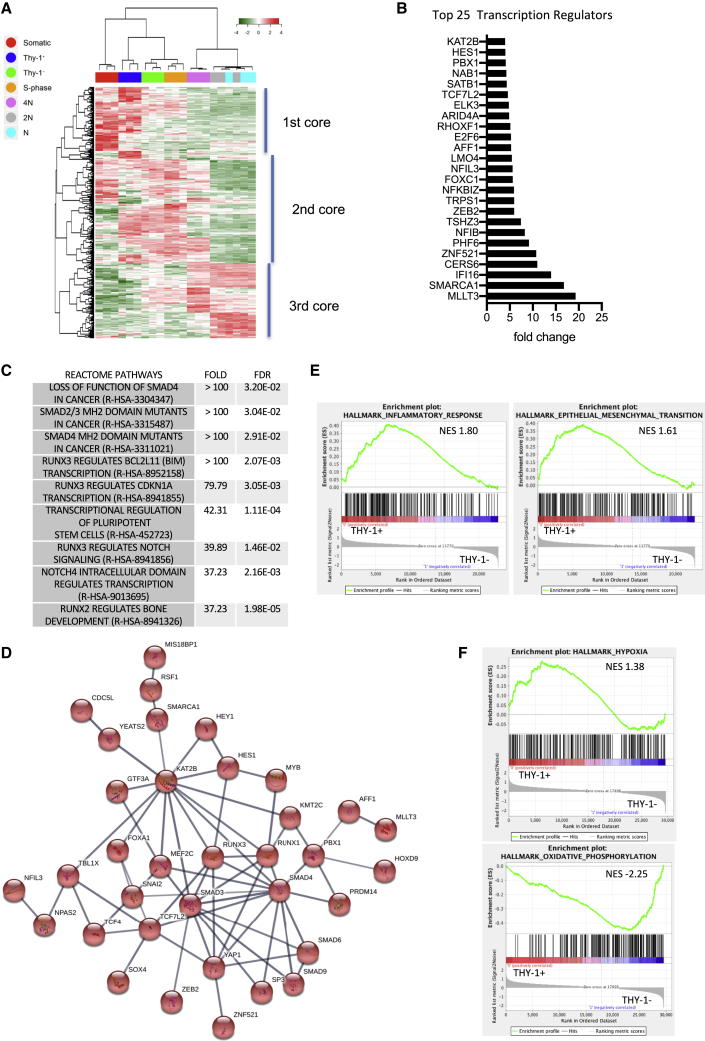
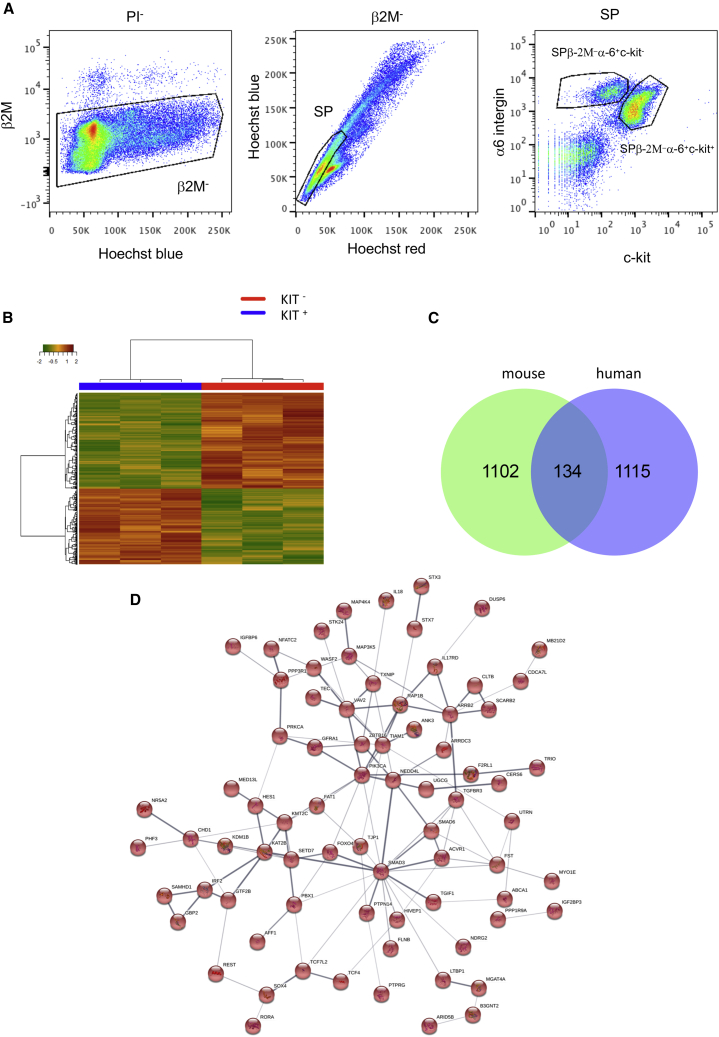
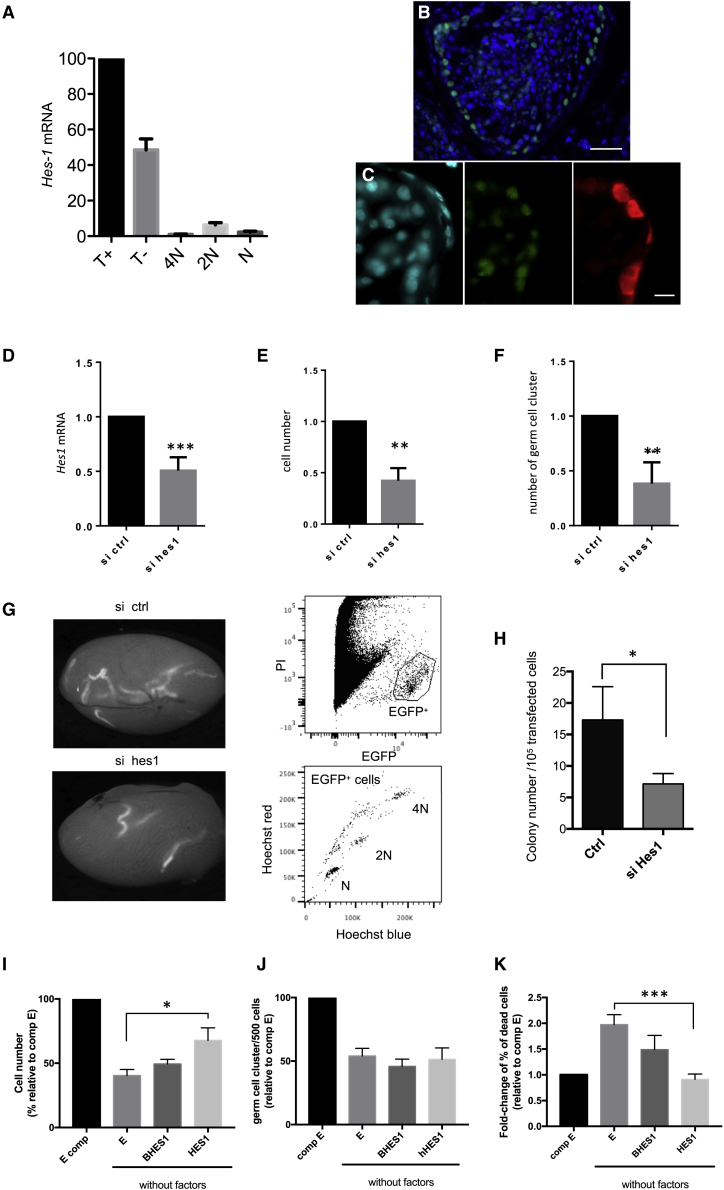
Male infertility is responsible for approximately half of all cases of reproductive issues. Spermatogenesis originates in a small pool of spermatogonial stem cells (SSCs), which are of interest for therapy of infertility but remain not well defined in humans. Using multiparametric analysis of the side population (SP) phenotype and the α-6 integrin, THY1, and β-2 microglobulin cell markers, we identified a population of human primitive undifferentiated spermatogonia with the phenotype β-2 microglobulin (β-2M)-SPα-6+THY1+, which is highly enriched in stem cells. By analyzing the expression signatures of this SSC-enriched population along with other germinal progenitors, we established an exhaustive transcriptome of human spermatogenesis. Transcriptome profiling of the human β-2M-SPα-6+THY1+ population and comparison with the profile of mouse undifferentiated spermatogonia provide insights into the molecular networks and key transcriptional regulators regulating human SSCs, including the basic-helix-loop-helix (bHLH) transcriptional repressor HES1, which we show to be implicated in maintenance of SSCs in vitro.
Keywords: human; spermatogenesis; stem cell; transcriptome.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Allen J.D., van Loevezijn A., Lakhai J.M., van der Valk M., van Tellingen O., Reid G., Schellens J.H., Koomen G.J., Schinkel A.H. Potent and specific inhibition of the breast cancer resistance protein multidrug transporter in vitro and in mouse intestine by a novel analogue of fumitremorgin C. Mol. Cancer Ther. 2002;1:417–425. - PubMed
-
- Barroca V., Lassalle B., Coureuil M., Louis J.P., Le Page F., Testart J., Allemand I., Riou L., Fouchet P. Mouse differentiating spermatogonia can generate germinal stem cells in vivo. Nat. Cell Biol. 2009;11:190–196. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
