

Structural basis of SARM1 activation, substrate recognition, and inhibition by small molecules
- PMID: 35334231
- PMCID: PMC9188649
- DOI: 10.1016/j.molcel.2022.03.007
Structural basis of SARM1 activation, substrate recognition, and inhibition by small molecules
Abstract
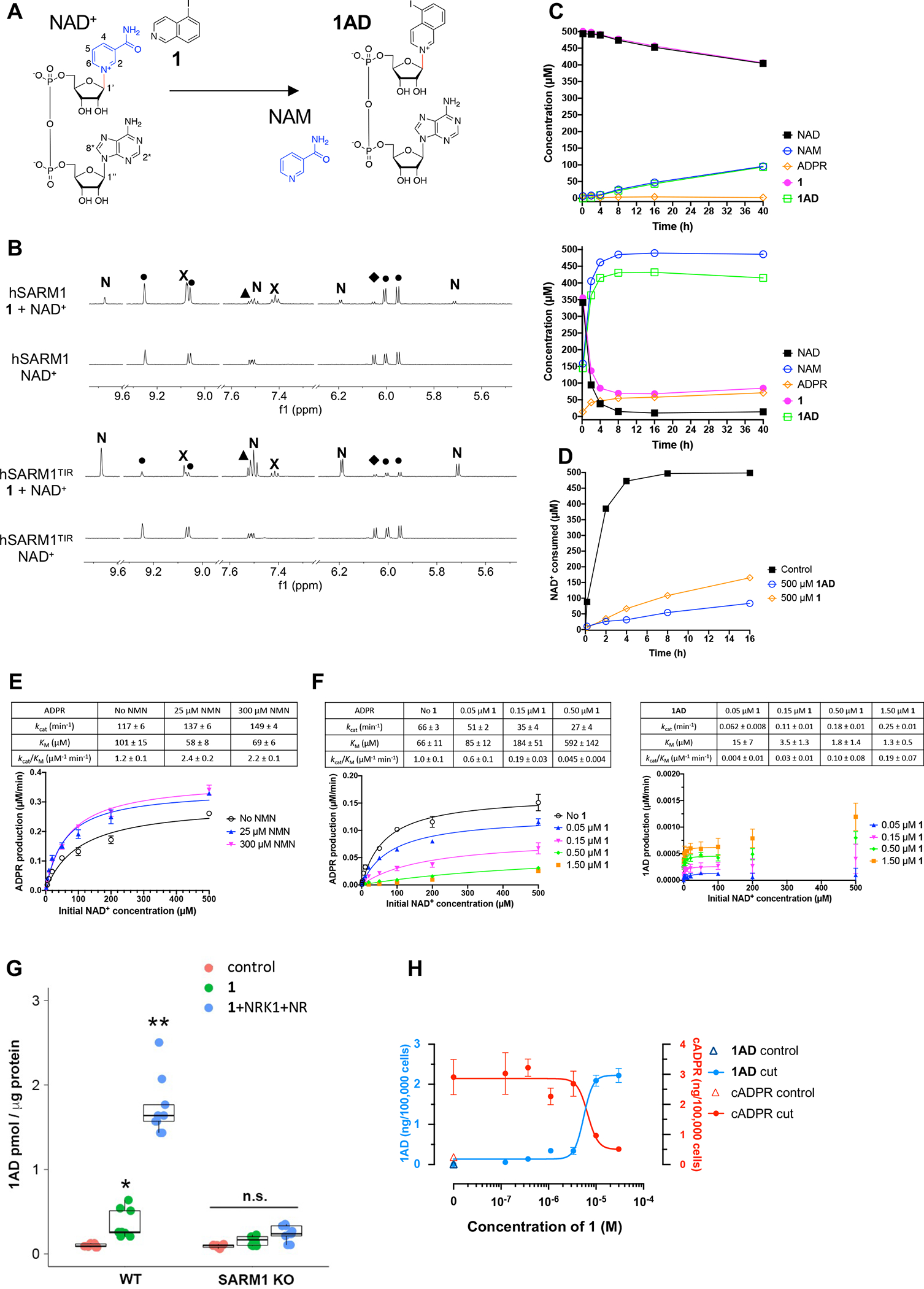
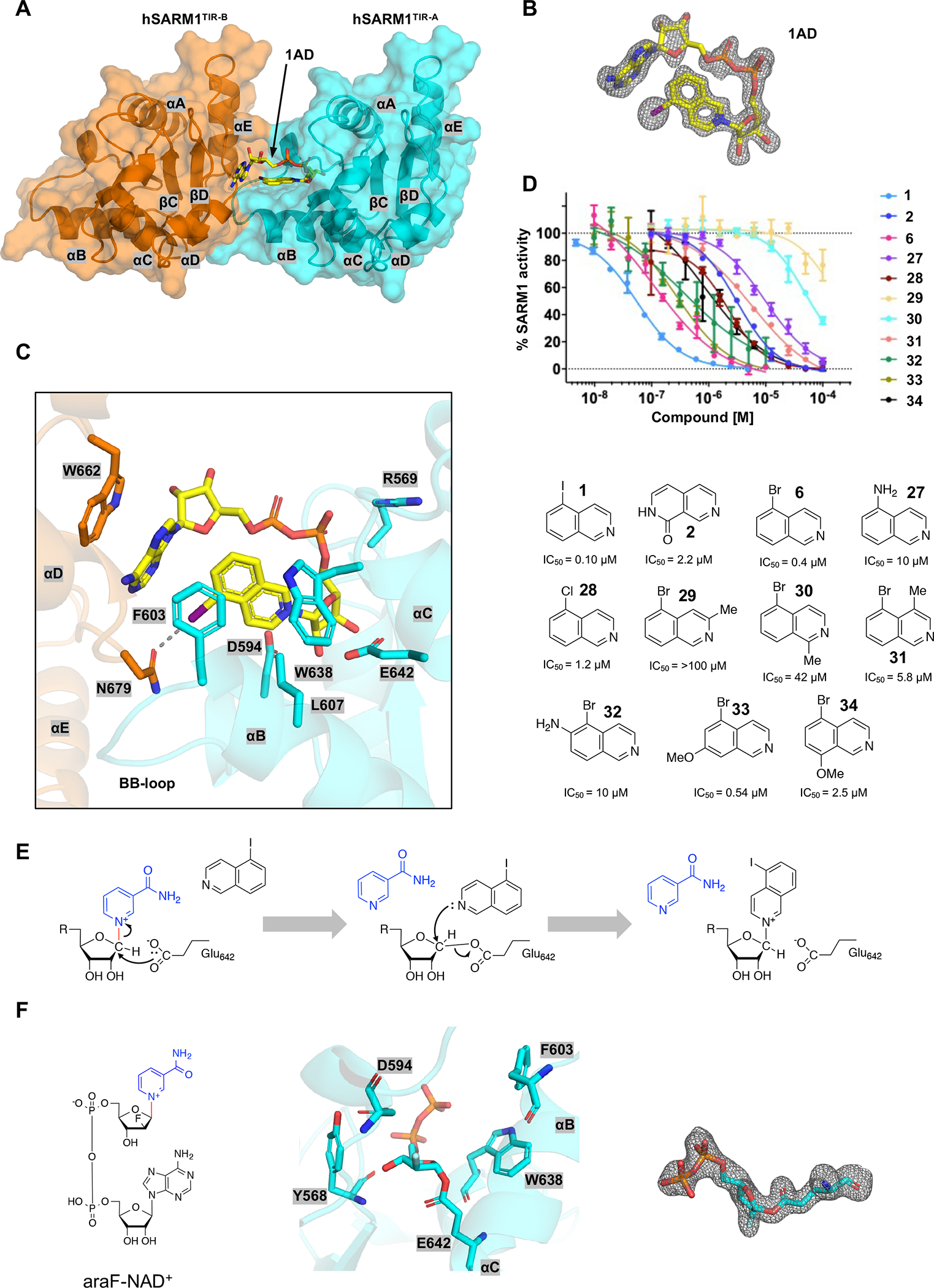
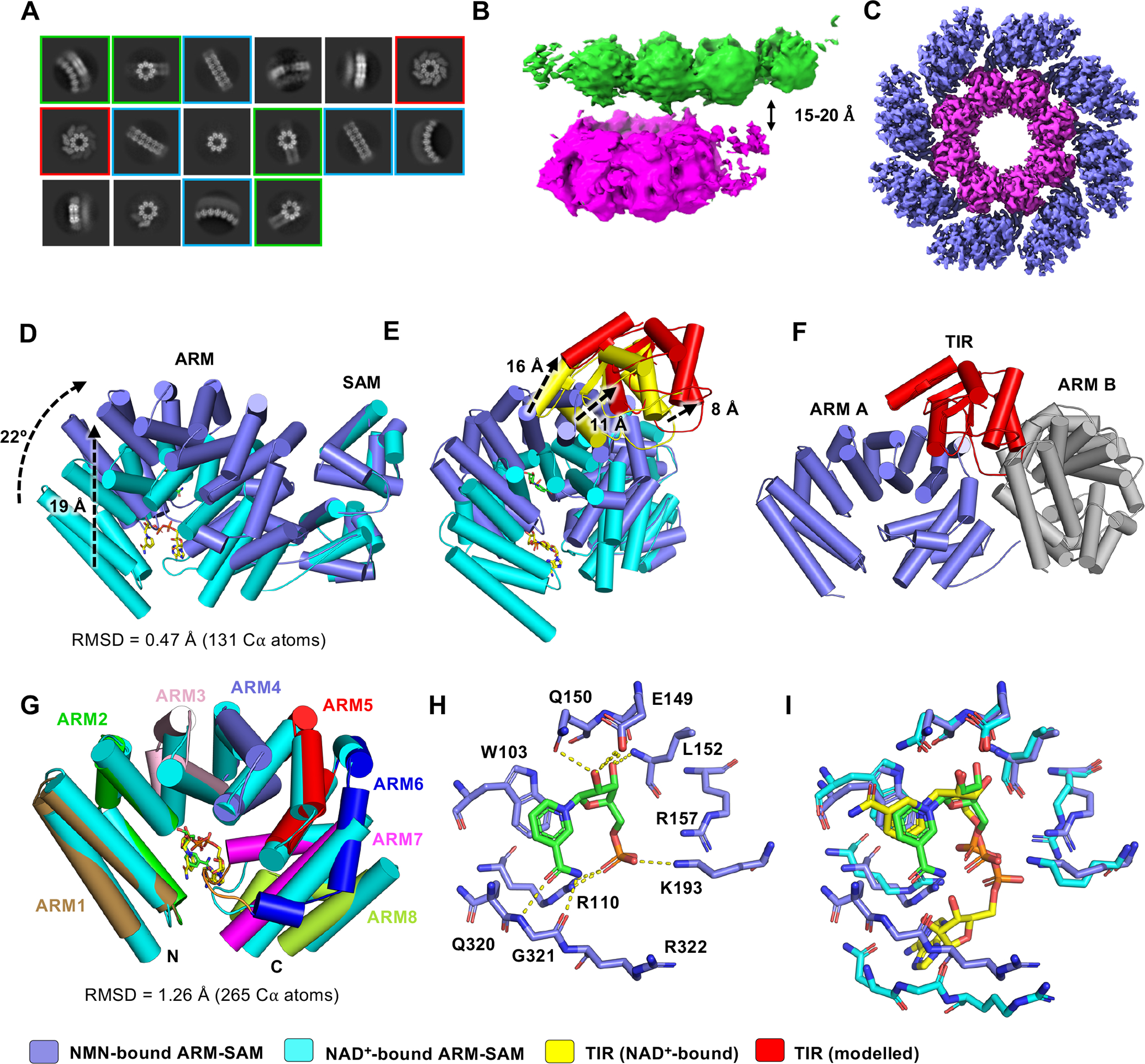
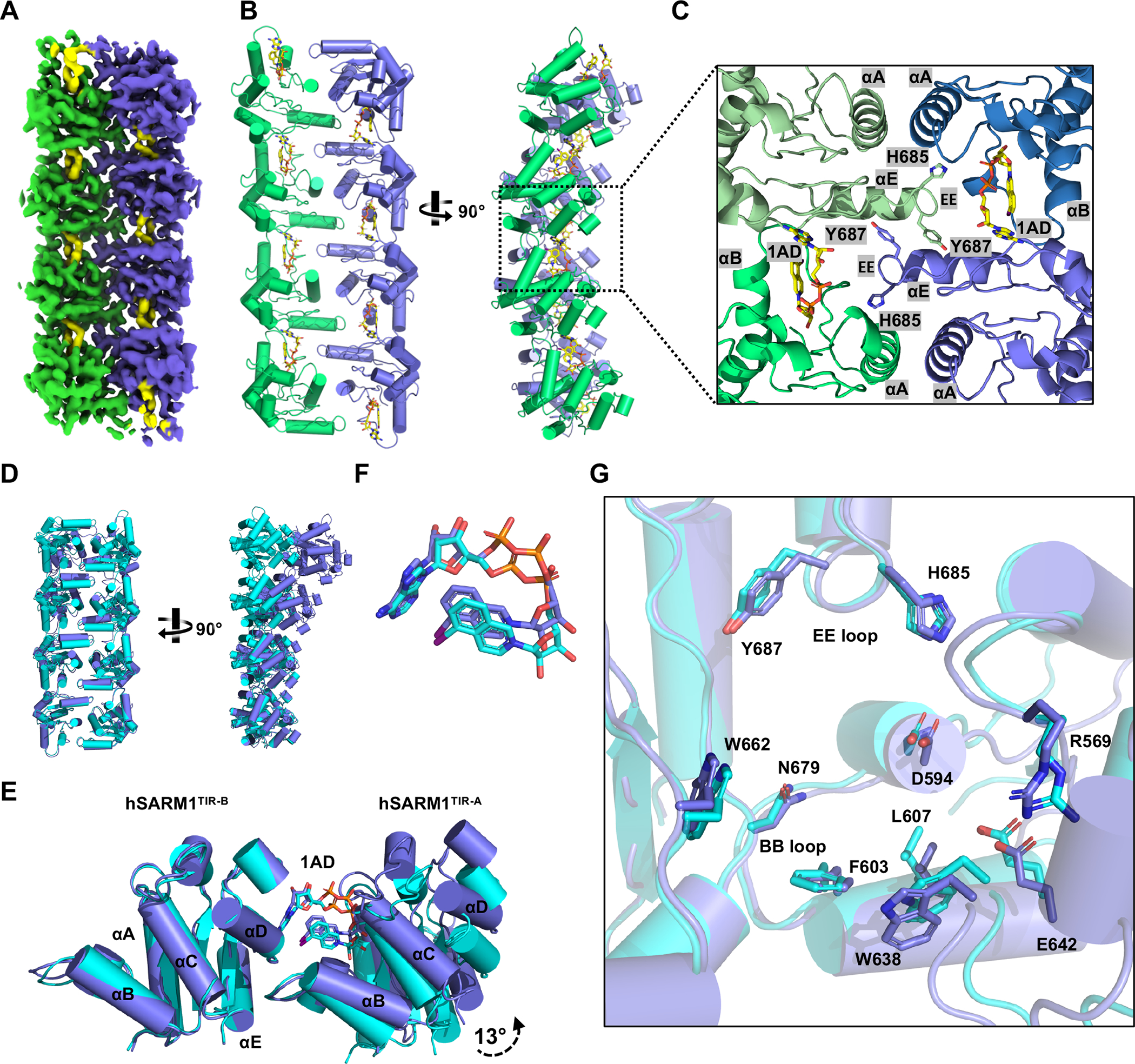
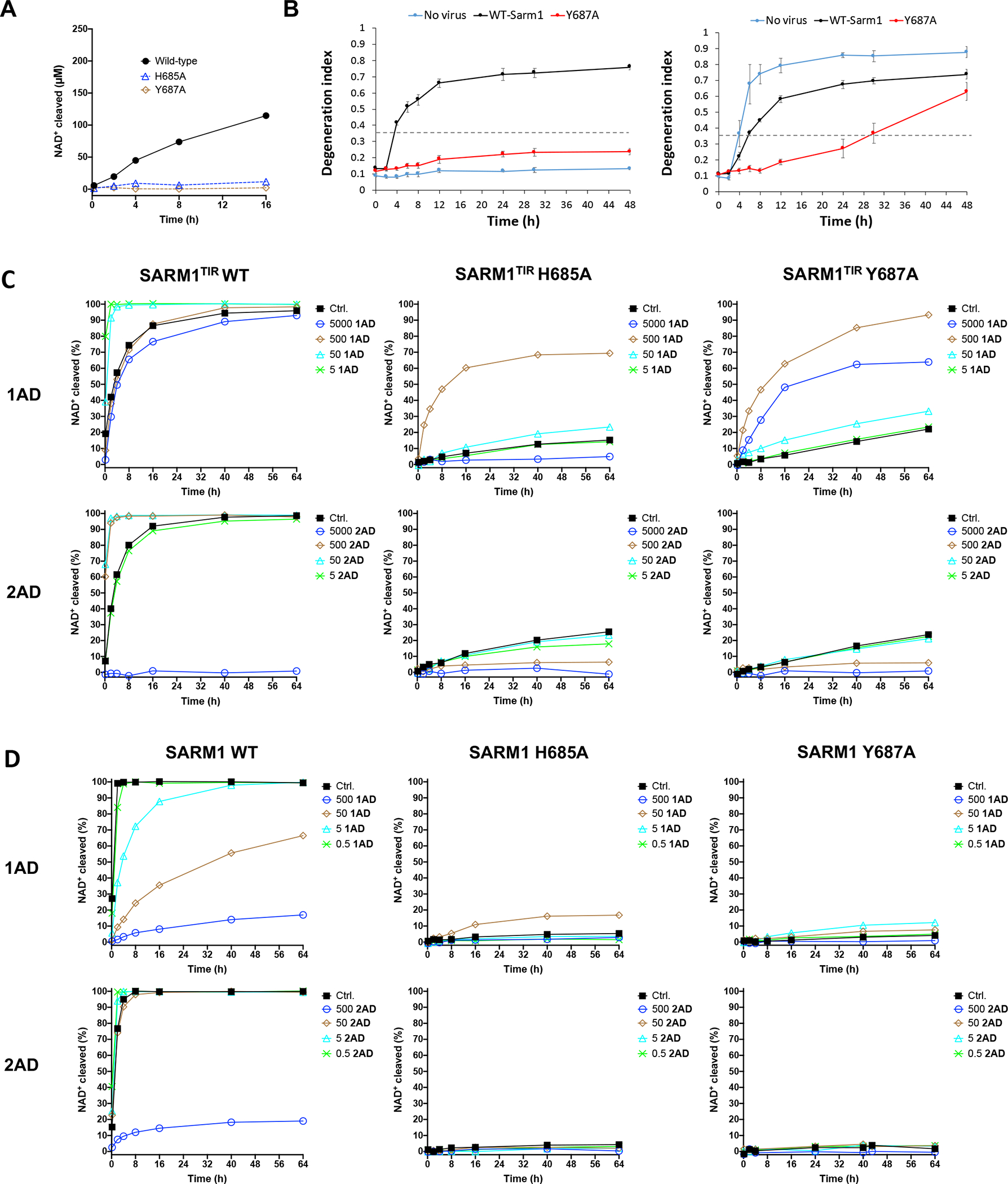
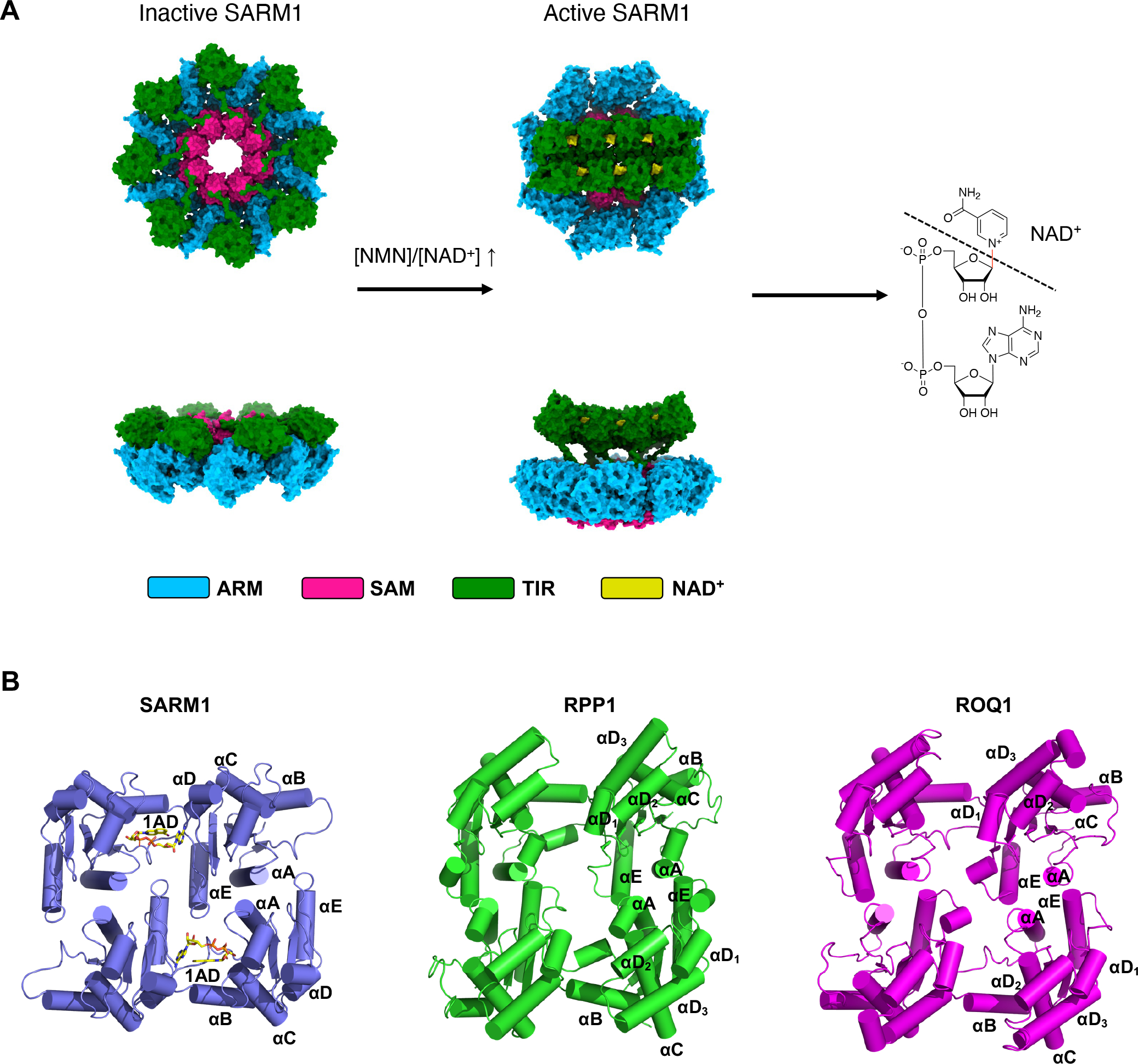
The NADase SARM1 (sterile alpha and TIR motif containing 1) is a key executioner of axon degeneration and a therapeutic target for several neurodegenerative conditions. We show that a potent SARM1 inhibitor undergoes base exchange with the nicotinamide moiety of nicotinamide adenine dinucleotide (NAD+) to produce the bona fide inhibitor 1AD. We report structures of SARM1 in complex with 1AD, NAD+ mimetics and the allosteric activator nicotinamide mononucleotide (NMN). NMN binding triggers reorientation of the armadillo repeat (ARM) domains, which disrupts ARM:TIR interactions and leads to formation of a two-stranded TIR domain assembly. The active site spans two molecules in these assemblies, explaining the requirement of TIR domain self-association for NADase activity and axon degeneration. Our results reveal the mechanisms of SARM1 activation and substrate binding, providing rational avenues for the design of new therapeutics targeting SARM1.
Keywords: ARM domain; NADase; TIR domain; X-ray crystallography; allosteric activator; base exchange; cryo-EM; orthosteric inhibitor.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.D. and J.M. are co-founders, scientific advisory board members, and shareholders of Disarm Therapeutics. B.K. is a shareholder of Disarm Therapeutics. Y. Sasaki and B.K. are consultants to Disarm Therapeutics. B.K. and T.V. receive research funding from Disarm Therapeutics. R.O.H., R.K., and T.B. are employees and shareholders in Disarm Therapeutics. S.E.A., M.F., K.C., A.B., and P.S.K. are employees of Evotec SE. R.O.H., T.B., and A.B. are inventors on a patent related to isoquinoline inhibitors of SARM1 (WO 2019/236879 Al). The authors have no additional competing financial interests.
Figures

References
-
- Bratkowski M, et al. (2020). Structural and mechanistic regulation of the pro-degenerative NAD hydrolase SARM1. Cell Rep 32, 107999. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
