

MicroRNA-200c Attenuates the Tumor-Infiltrating Capacity of Macrophages
- PMID: 35336722
- PMCID: PMC8945044
- DOI: 10.3390/biology11030349
MicroRNA-200c Attenuates the Tumor-Infiltrating Capacity of Macrophages
Abstract
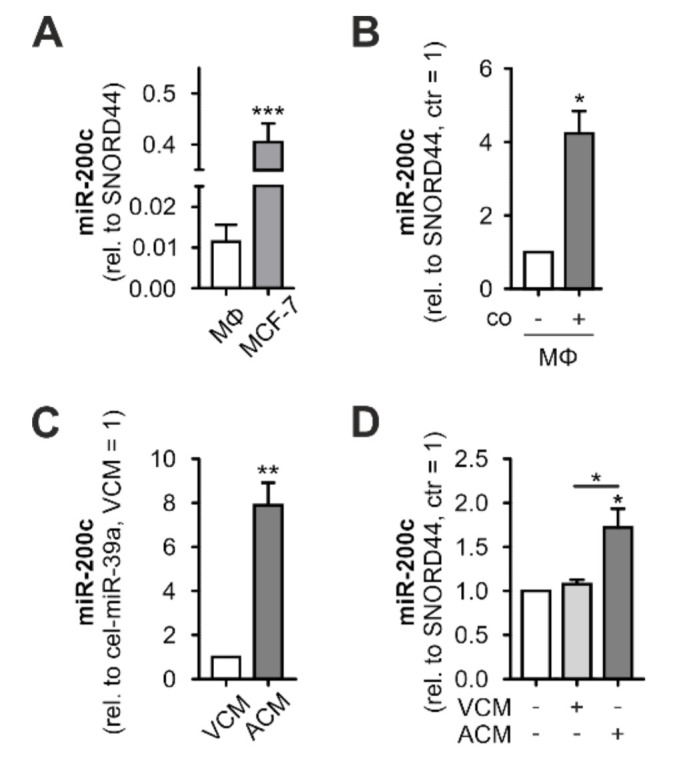
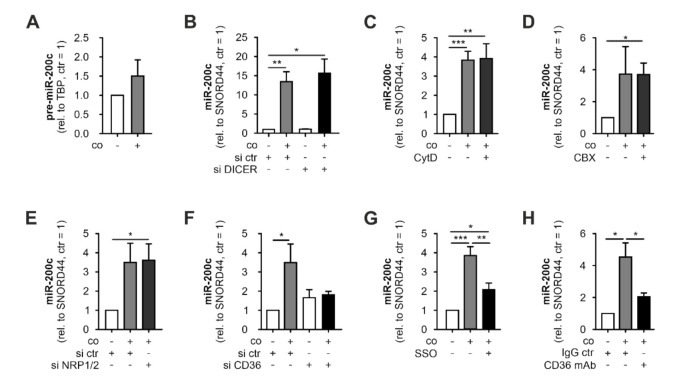
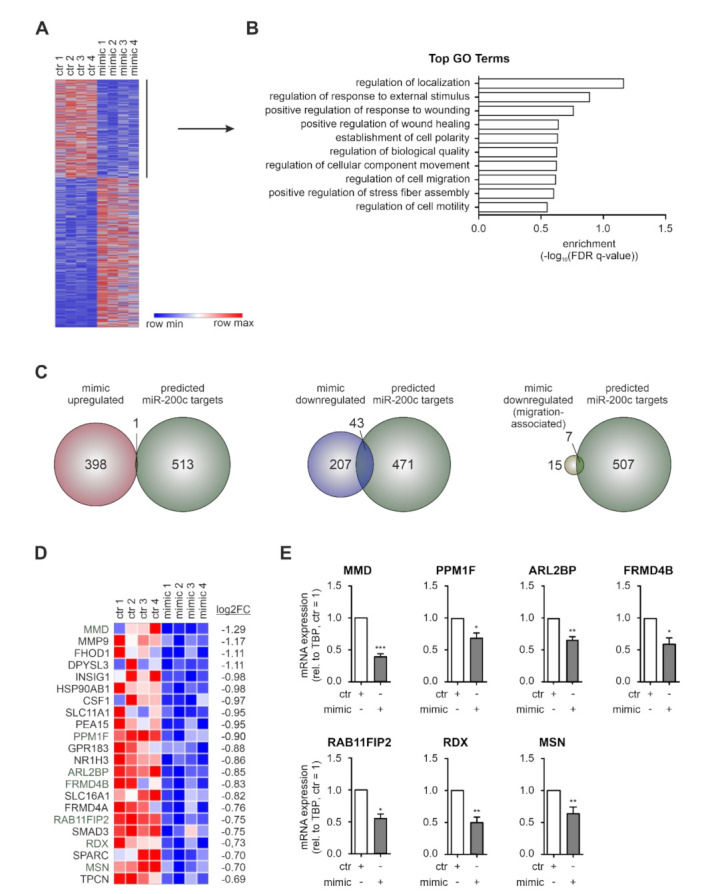
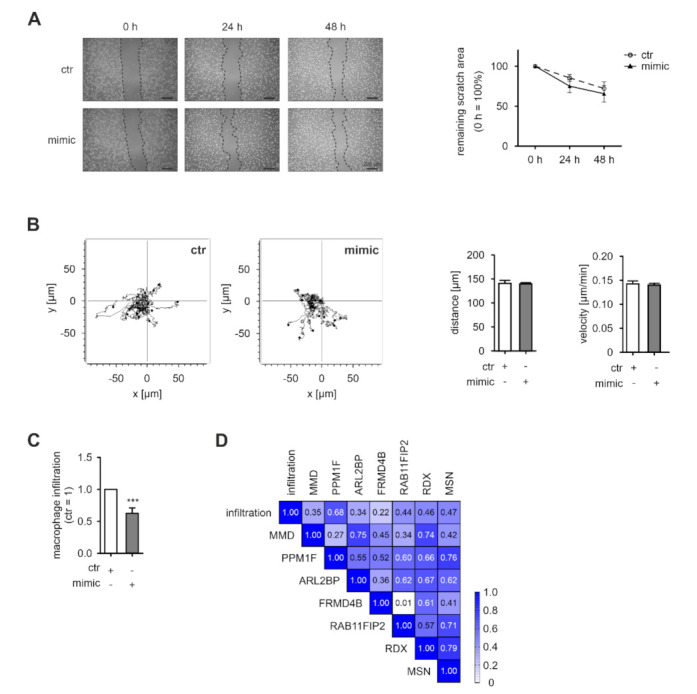
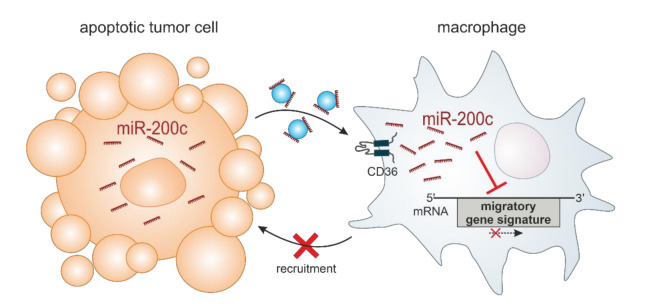
Macrophages constitute a major part of the tumor-infiltrating immune cells. Within the tumor microenvironment, they acquire an alternatively activated, tumor-supporting phenotype. Factors released by tumor cells are crucial for the recruitment of tumor-associated macrophages. In the present project, we aimed to understand the role of hsa-miR-200c-3p (miR-200c) in the interplay between tumor cells and macrophages. To this end, we employed a coculture system of MCF7 breast tumor cells and primary human macrophages and observed the transfer of miR-200c from apoptotic tumor cells to macrophages, which required intact CD36 receptor in macrophages. We further comprehensively determined miR-200c targets in macrophages by mRNA-sequencing and identified numerous migration-associated mRNAs to be downregulated by miR-200c. Consequently, miR-200c attenuated macrophage infiltration into 3-dimensional tumor spheroids. miR-200c-mediated reduction in infiltration further correlated with a miR-200c migration signature comprised of the four miR-200c-repressed, predicted targets PPM1F, RAB11FIB2, RDX, and MSN.
Keywords: breast tumor; macrophage; miR; tumor microenvironment.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- World Health Organisation Breast Cancer. [(accessed on 8 September 2021)]. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer.
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
