

Five decades of misunderstanding in the social Hymenoptera: a review and meta-analysis of Michener's paradox
- PMID: 35338566
- PMCID: PMC9546470
- DOI: 10.1111/brv.12854
Five decades of misunderstanding in the social Hymenoptera: a review and meta-analysis of Michener's paradox
Abstract
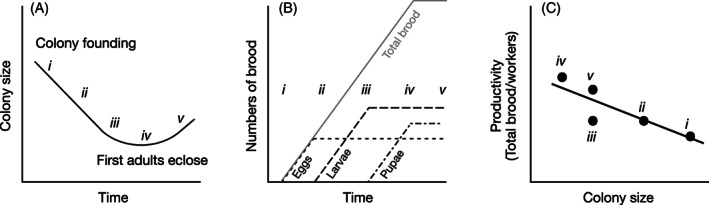
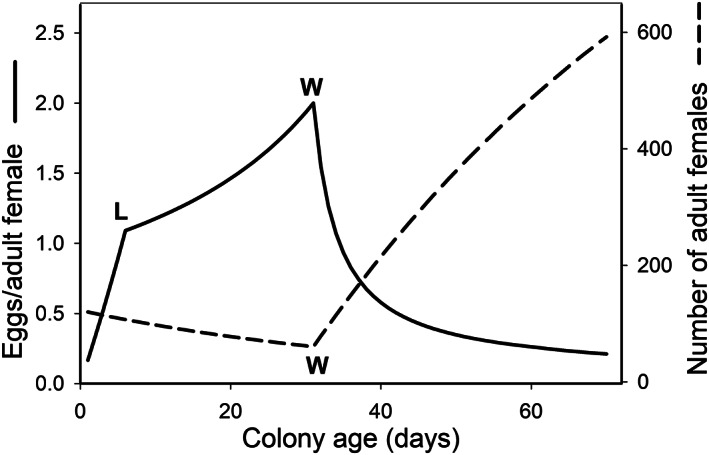
In a much-cited 1964 paper entitled "Reproductive efficiency in relation to colony size in hymenopterous societies," Charles Michener investigated the correlation between a colony's size and its reproductive efficiency - the ability of its adult females to produce reproductives, measured as per-capita output. Based on his analysis of published data from destructively sampled colonies in 18 species, he reported that in most of these species efficiency decreased with increasing colony size. His conclusion that efficiency is higher in smaller groups has since gained widespread acceptance. But it created a seeming paradox: how can natural selection maintain social behaviour when a female apparently enjoys her highest per-capita output by working alone? Here we treat Michener's pattern as a hypothesis and perform the first large-scale test of its prediction across the eusocial Hymenoptera. Because data on actual output of reproductives were not available for most species, Michener used various proxies, such as nest size, numbers of brood, or amounts of stored food. We show that for each of Michener's data sets the reported decline in per-capita productivity can be explained by factors other than decreasing efficiency, calling into question his conclusion that declining efficiency is the cause of the pattern. The most prominent cause of bias is the failure of the proxy to capture all forms of output in which the colony invests during the course of its ontogeny. Other biasing factors include seasonal effects and a variety of methodological flaws in the data sets he used. We then summarize the results of 215 data sets drawn from post-1964 studies of 80 species in 33 genera that better control for these factors. Of these, 163 data sets are included in two meta-analyses that statistically synthesize the available data on the relationship between colony size and efficiency, accounting for variable sample sizes and non-independence among the data sets. The overall effect, and those for most taxonomic subgroups, indicates no loss of efficiency with increasing colony size. Two exceptional taxa, the halictid bees and independent-founding paper wasps, show negative trends consistent with the Michener hypothesis in some species. We conclude that in most species, particularly those with large colony sizes, the hypothesis of decreasing efficiency with increasing colony size is not supported. Finally, we explore potential mechanisms through which the level of efficiency can decrease, be maintained, or even increase, as colonies increase in size.
Keywords: Hymenoptera; colony development; colony size; division of labour; ergonomic efficiency; social evolution.
© 2022 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures









References
-
- Accorti, M. , Tarducci, F. & Luti, F. (1991). Numerical ratio between adult bees and reared brood. Ethology Ecology & Evolution 3(sup1), 119–122.
-
- Adams, D. C. (2008). Phylogenetic meta‐analysis. Evolution 62(3), 567–572. - PubMed
-
- Akre, R. D. , Garnett, W. B. , Mac Donald, J. F. , Greene, A. & Landolt, P. (1976). Behavior and colony development of Vespula pensylvanica and V. atropilosa (Hymenoptera: Vespidae). Journal of the Kansas Entomological Society 49(1), 63–84.
-
- Anderson, C. , Boomsma, J. J. & Bartholdi, J. J. (2002). Task partitioning in insect societies: bucket brigades. Insectes Sociaux 49(2), 171–180.
-
- Anderson, C. & Franks, N. (2001). Teams in animal societies. Behavioral Ecology 12(5), 534–540.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
