

Acquired Disorder and Asymmetry in a Domain-Swapped Model for γ-Crystallin Aggregation
- PMID: 35341744
- PMCID: PMC9050881
- DOI: 10.1016/j.jmb.2022.167559
Acquired Disorder and Asymmetry in a Domain-Swapped Model for γ-Crystallin Aggregation
Abstract
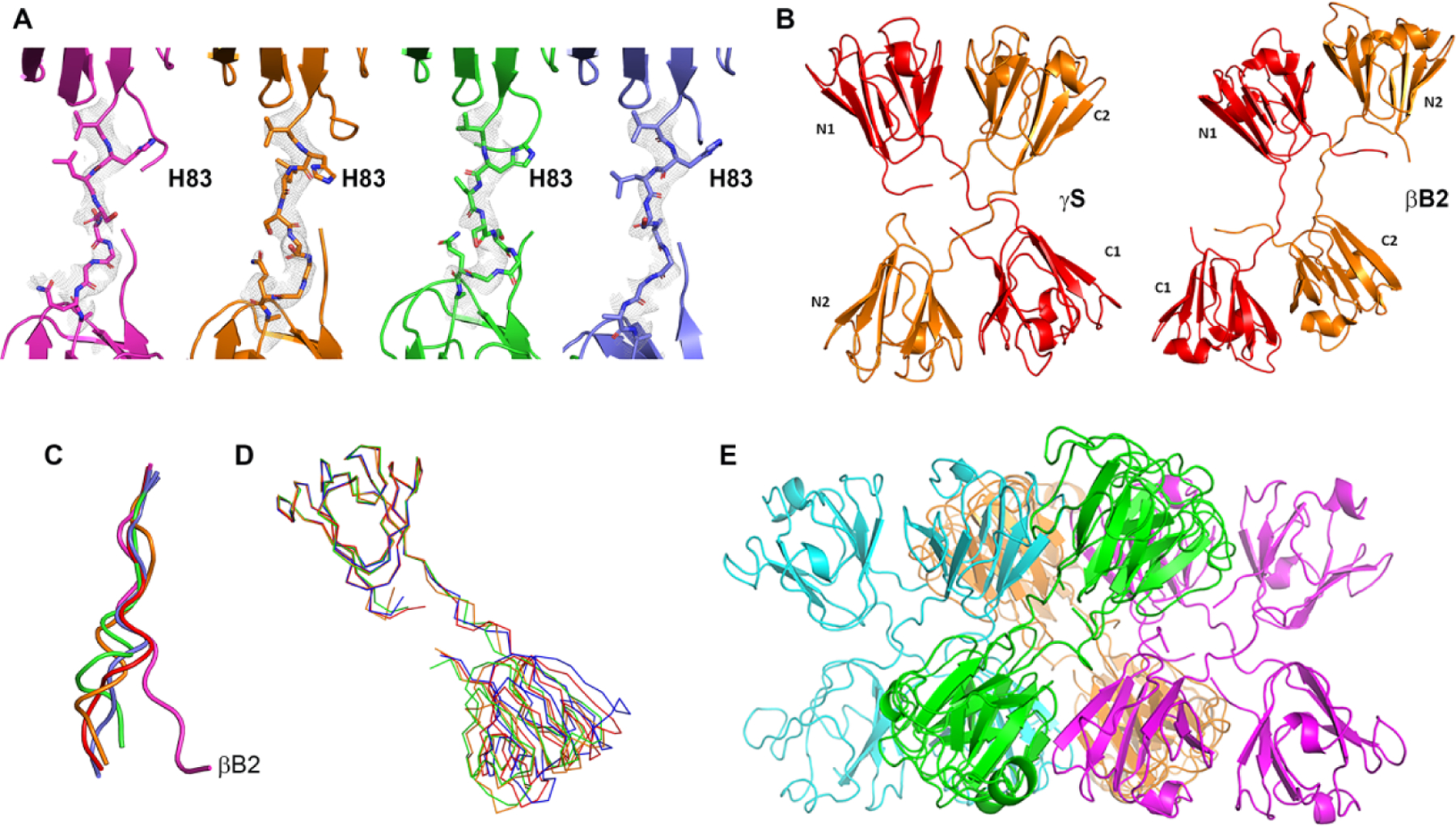
Misfolding and aggregation of proteins occur in many pathological states. Because of the inherent disorder involved, these processes are difficult to study. We attempted to capture aggregation intermediates of γS-crystallin, a highly stable, internally symmetrical monomeric protein, by crystallization under mildly acidic and oxidizing conditions. Here we describe novel oligomerization through strained domain-swapping and partial intermolecular disulfide formation. This forms an octamer built from asymmetric tetramers, each of which comprises an asymmetric pair of twisted, domain-swapped dimers. Each tetramer shows patterns of acquired disorder among subunits, ranging from local loss of secondary structure to regions of intrinsic disorder. The octamer ring is tied together by partial intermolecular disulfide bonds, which may contribute to strain and disorder in the octamer. Oligomerization in this structure is self-limited by the distorted octamer ring. In a more heterogeneous environment, the disordered regions could serve as seeds for cascading interactions with other proteins. Indeed, solubilized protein from crystals retain many features observed in the crystal and are prone to further oligomerization and precipitation. This structure illustrates modes of loss of organized structure and aggregation that are relevant for cataract and for other disorders involving deposition of formerly well-folded proteins.
Keywords: aggregation; asymmetry; disorder; domain-swapping; unfolding.
Published by Elsevier Ltd.
Conflict of interest statement
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures





References
-
- Chiti F & Dobson CM (2006). Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem 75, 333–66. - PubMed
-
- Dyson HJ & Wright PE (2005). Intrinsically unstructured proteins and their functions. Nat Rev Mol Cell Biol 6, 197–208. - PubMed
-
- Ferina J & Daggett V (2019). Visualizing Protein Folding and Unfolding. J Mol Biol 431, 1540–1564. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
