

Genome-wide CRISPR screen reveals CLPTM1L as a lipid scramblase required for efficient glycosylphosphatidylinositol biosynthesis
- PMID: 35344438
- PMCID: PMC9169118
- DOI: 10.1073/pnas.2115083119
Genome-wide CRISPR screen reveals CLPTM1L as a lipid scramblase required for efficient glycosylphosphatidylinositol biosynthesis
Abstract
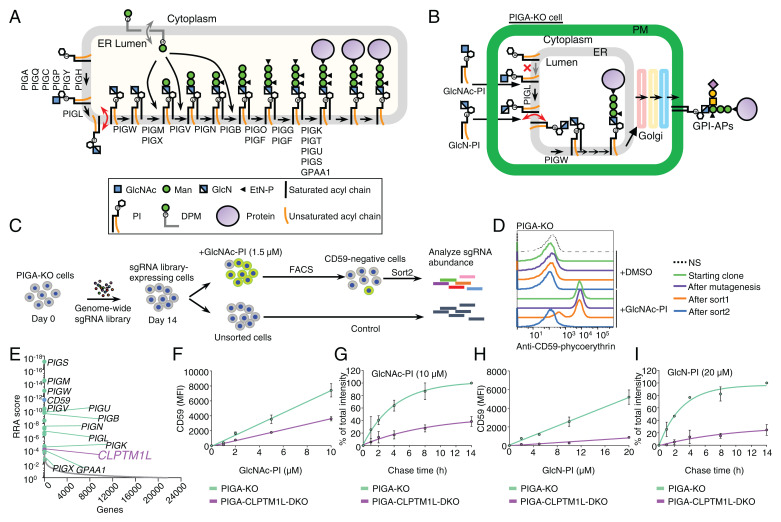
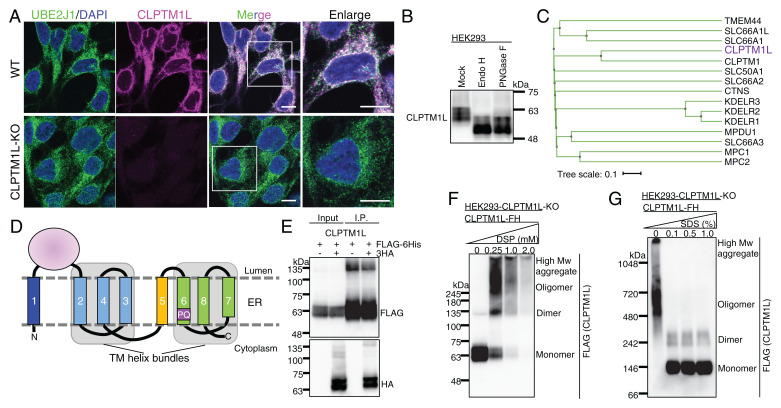
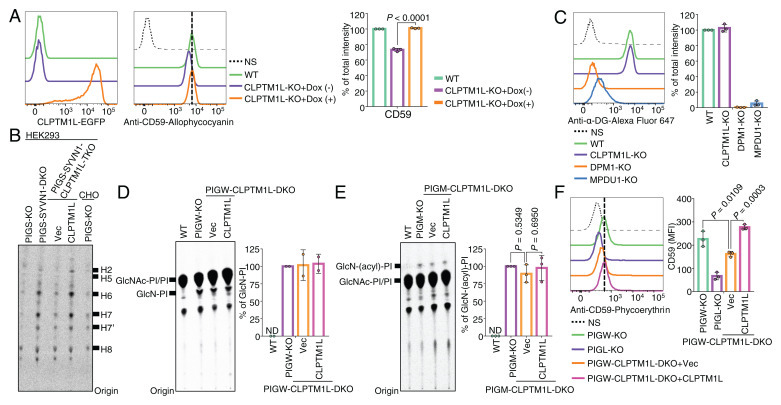
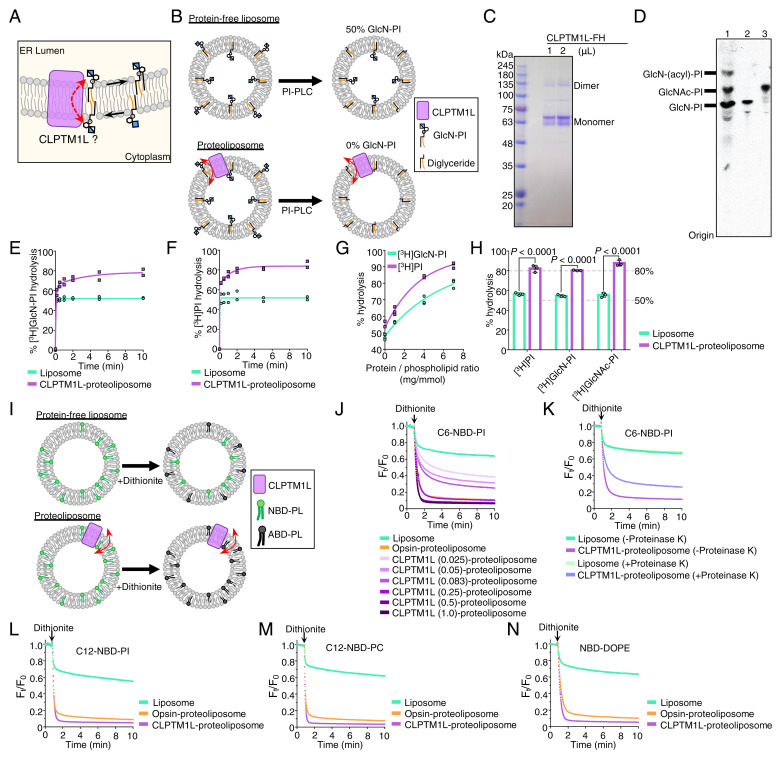
SignificanceScramblases translocate lipids across the lipid bilayer without consumption of ATP, thereby regulating lipid distributions in cellular membranes. Cytosol-to-lumen translocation across the endoplasmic reticulum (ER) membrane is a common process among lipid glycoconjugates involved in posttranslational protein modifications in eukaryotes. These translocations are thought to be mediated by specific ER-resident scramblases, but the identity of these proteins and the underlying molecular mechanisms have been elusive. Here, we show that CLPTM1L, an integral membrane protein with eight putative transmembrane domains, is the major lipid scramblase involved in efficient glycosylphosphatidylinositol biosynthesis in the ER membrane. Our results validate the long-standing hypothesis that lipid scramblases ensure the efficient translocations of lipid glycoconjugates across the ER membrane for protein glycosylation pathways.
Keywords: CLPTM1L; endoplasmic reticulum; glycobiology; glycosylphosphatidylinositol; scramblase.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Two endoplasmic reticulum (ER) membrane proteins that facilitate ER-to-Golgi transport of glycosylphosphatidylinositol-anchored proteins.Mol Biol Cell. 1999 Apr;10(4):1043-59. doi: 10.1091/mbc.10.4.1043. Mol Biol Cell. 1999. PMID: 10198056 Free PMC article.
-
TMEM41B acts as an ER scramblase required for lipoprotein biogenesis and lipid homeostasis.Cell Metab. 2021 Aug 3;33(8):1655-1670.e8. doi: 10.1016/j.cmet.2021.05.006. Epub 2021 May 19. Cell Metab. 2021. PMID: 34015269
-
Trafficking of glycosylphosphatidylinositol anchored proteins from the endoplasmic reticulum to the cell surface.J Lipid Res. 2016 Mar;57(3):352-60. doi: 10.1194/jlr.R062760. Epub 2015 Oct 8. J Lipid Res. 2016. PMID: 26450970 Free PMC article. Review.
-
Yeast ARV1 is required for efficient delivery of an early GPI intermediate to the first mannosyltransferase during GPI assembly and controls lipid flow from the endoplasmic reticulum.Mol Biol Cell. 2008 May;19(5):2069-82. doi: 10.1091/mbc.e07-08-0740. Epub 2008 Feb 20. Mol Biol Cell. 2008. PMID: 18287539 Free PMC article.
-
Transport of glycosylphosphatidylinositol-anchored proteins from the endoplasmic reticulum.Biochim Biophys Acta. 2013 Nov;1833(11):2473-8. doi: 10.1016/j.bbamcr.2013.01.027. Epub 2013 Feb 1. Biochim Biophys Acta. 2013. PMID: 23380706 Review.
Cited by
-
Multi-omic characterization of the maize GPI synthesis mutant gwt1 with defects in kernel development.BMC Plant Biol. 2023 Apr 10;23(1):191. doi: 10.1186/s12870-023-04188-w. BMC Plant Biol. 2023. PMID: 37038106 Free PMC article.
-
Long-term benefits of TUDCA supplement in ARSACS zebrafish model.Sci Rep. 2025 Jul 14;15(1):25429. doi: 10.1038/s41598-025-10850-0. Sci Rep. 2025. PMID: 40659819 Free PMC article.
-
Recent research progress in glycosylphosphatidylinositol-anchored protein biosynthesis, chemical/chemoenzymatic synthesis, and interaction with the cell membrane.Curr Opin Chem Biol. 2024 Feb;78:102421. doi: 10.1016/j.cbpa.2023.102421. Epub 2024 Jan 4. Curr Opin Chem Biol. 2024. PMID: 38181647 Free PMC article. Review.
-
Inherited glycosylphosphatidylinositol deficiency: a review from molecular and clinical perspectives.Acta Biochim Biophys Sin (Shanghai). 2024 Jul 30;56(8):1234-1243. doi: 10.3724/abbs.2024128. Acta Biochim Biophys Sin (Shanghai). 2024. PMID: 39081219 Free PMC article. Review.
-
Parallel CRISPR-Cas9 screens identify mechanisms of PLIN2 and lipid droplet regulation.Dev Cell. 2023 Sep 25;58(18):1782-1800.e10. doi: 10.1016/j.devcel.2023.07.001. Epub 2023 Jul 25. Dev Cell. 2023. PMID: 37494933 Free PMC article.
References
-
- Takeda J., et al. , Deficiency of the GPI anchor caused by a somatic mutation of the PIG-A gene in paroxysmal nocturnal hemoglobinuria. Cell 73, 703–711 (1993). - PubMed
-
- Bellai-Dussault K., Nguyen T. T. M., Baratang N. V., Jimenez-Cruz D. A., Campeau P. M., Clinical variability in inherited glycosylphosphatidylinositol deficiency disorders. Clin. Genet. 95, 112–121 (2019). - PubMed
-
- Pottekat A., Menon A. K., Subcellular localization and targeting of N-acetylglucosaminyl phosphatidylinositol de-N-acetylase, the second enzyme in the glycosylphosphatidylinositol biosynthetic pathway. J. Biol. Chem. 279, 15743–15751 (2004). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
