

Aquatic birds have middle ears adapted to amphibious lifestyles
- PMID: 35347167
- PMCID: PMC8960762
- DOI: 10.1038/s41598-022-09090-3
Aquatic birds have middle ears adapted to amphibious lifestyles
Abstract
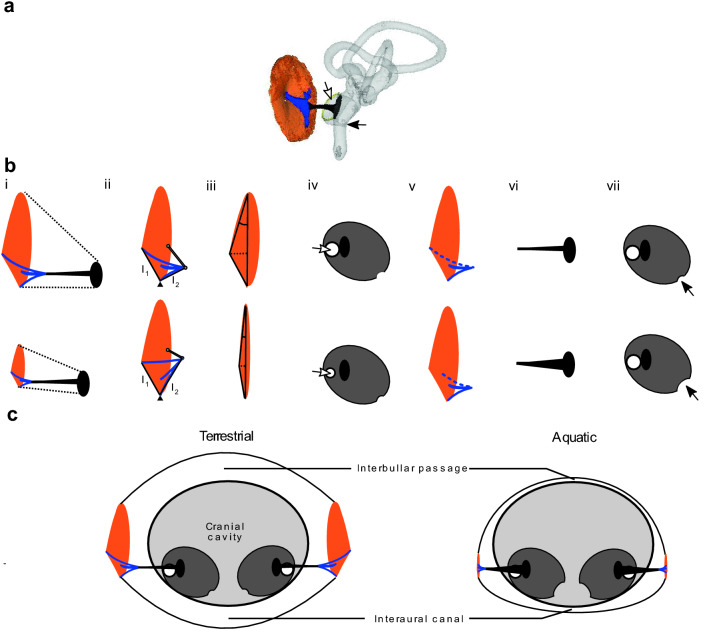
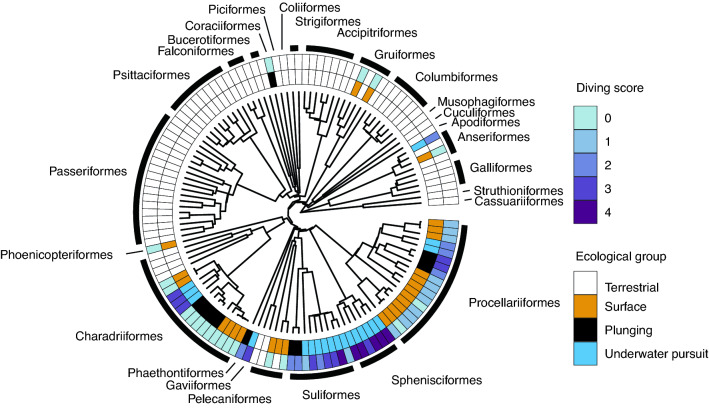
Birds exhibit wide variation in their use of aquatic environments, on a spectrum from entirely terrestrial, through amphibious, to highly aquatic. Although there are limited empirical data on hearing sensitivity of birds underwater, mounting evidence indicates that diving birds detect and respond to sound underwater, suggesting that some modifications of the ear may assist foraging or other behaviors below the surface. In air, the tympanic middle ear acts as an impedance matcher that increases sound pressure and decreases sound vibration velocity between the outside air and the inner ear. Underwater, the impedance-matching task is reversed and the ear is exposed to high hydrostatic pressures. Using micro- and nano-CT (computerized tomography) scans of bird ears in 127 species across 26 taxonomic orders, we measured a suite of morphological traits of importance to aerial and aquatic hearing to test predictions relating to impedance-matching in birds with distinct aquatic lifestyles, while accounting for allometry and phylogeny. Birds that engage in underwater pursuit and deep diving showed the greatest differences in ear structure relative to terrestrial species. In these heavily modified ears, the size of the input areas of both the tympanic membrane and the columella footplate of the middle ear were reduced. Underwater pursuit and diving birds also typically had a shorter extrastapedius, a reduced cranial air volume and connectivity and several modifications in line with reversals of low-to-high impedance-matching. The results confirm adaptations of the middle ear to aquatic lifestyles in multiple independent bird lineages, likely facilitating hearing underwater and baroprotection, while potentially constraining the sensitivity of aerial hearing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Saunders JC, Duncan RK, Doan DE, Werner YL. The middle ear of reptiles and birds. In: Dooling RJ, Fay RR, Popper AN, editors. Comparative Hearing: Birds and Reptiles. Springer; 2000. pp. 13–69.
-
- Christensen-Dalsgaard J, Manley GA. The malleable middle ear: An underappreciated player in the evolution of hearing in vertebrates. In: Köppl C, Manley GA, Popper AN, Fay RR, editors. Insights from Comparative Hearing Research. Springer; 2013. pp. 157–191.
-
- Hemilä S, Nummela S, Reuter T. A model of the odontocete middle ear. Hear. Res. 1999;133:82–97. - PubMed
-
- Nummela S, Thewissen JGM, Bajpai S, Hussain T, Kumar K. Sound transmission in archaic and modern whales: Anatomical adaptations for underwater hearing. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2007;290:716–733. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
