

Adenosine signaling activates ATP-sensitive K+ channels in endothelial cells and pericytes in CNS capillaries
- PMID: 35349300
- PMCID: PMC9623876
- DOI: 10.1126/scisignal.abl5405
Adenosine signaling activates ATP-sensitive K+ channels in endothelial cells and pericytes in CNS capillaries
Abstract
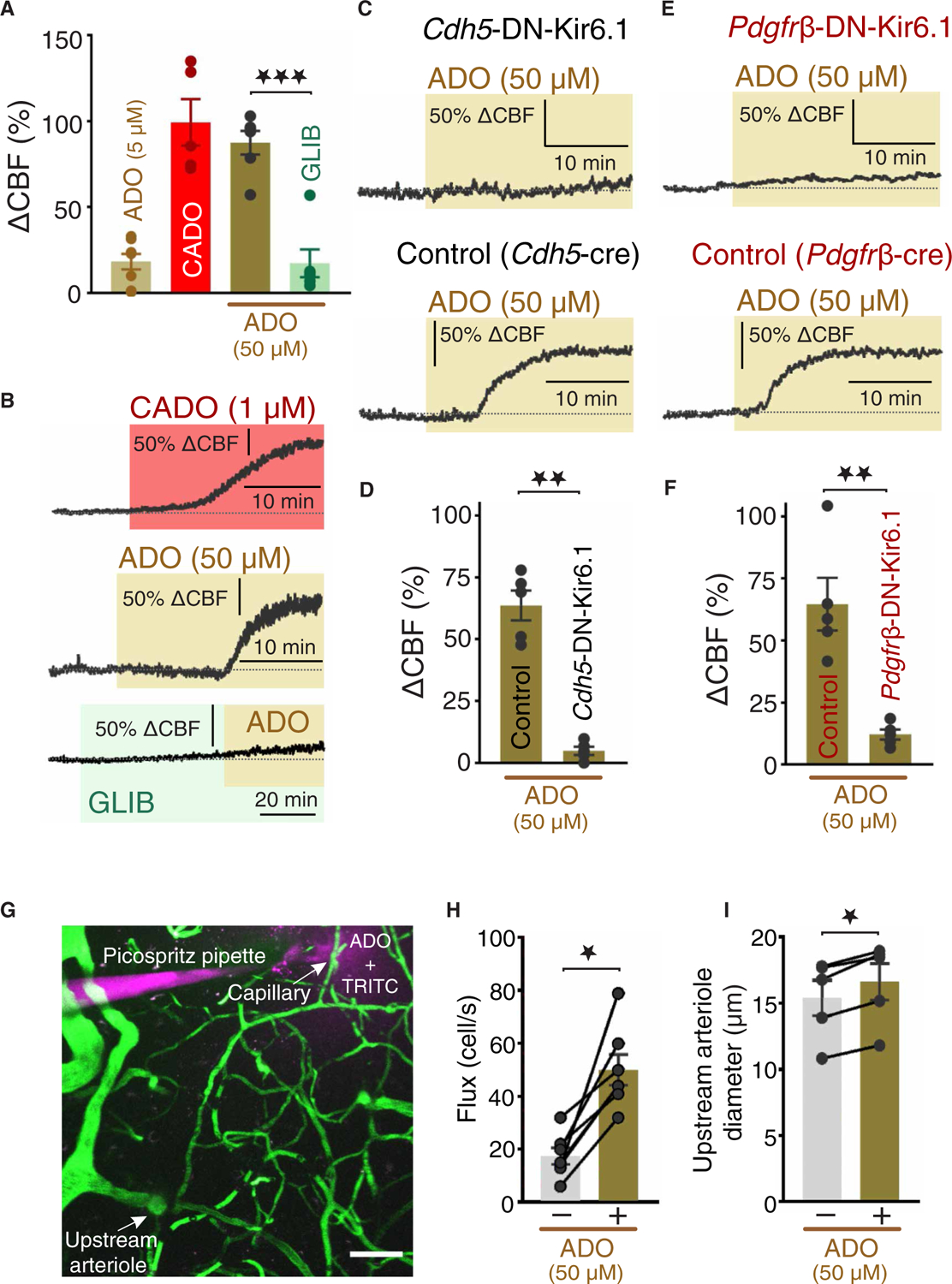
The dense network of capillaries composed of capillary endothelial cells (cECs) and pericytes lies in close proximity to all neurons, ideally positioning it to sense neuron- and glial-derived compounds that enhance regional and global cerebral perfusion. The membrane potential (VM) of vascular cells serves as the physiological bridge that translates brain activity into vascular function. In other beds, the ATP-sensitive K+ (KATP) channel regulates VM in vascular smooth muscle, which is absent in the capillary network. Here, with transgenic mice that expressed a dominant-negative mutant of the pore-forming Kir6.1 subunit specifically in brain cECs or pericytes, we demonstrated that KATP channels were present in both cell types and robustly controlled VM. We further showed that the signaling nucleotide adenosine acted through A2A receptors and the Gαs/cAMP/PKA pathway to activate capillary KATP channels. Moreover, KATP channel stimulation in vivo increased cerebral blood flow (CBF), an effect that was blunted by expression of the dominant-negative Kir6.1 mutant in either capillary cell type. These findings establish an important role for KATP channels in cECs and pericytes in the regulation of CBF.
Conflict of interest statement
Figures






Comment in
-
Tuning the signal: ATP-sensitive K+ channels direct blood flow to cerebral capillaries.Sci Signal. 2022 Mar 29;15(727):eabo1118. doi: 10.1126/scisignal.abo1118. Epub 2022 Mar 29. Sci Signal. 2022. PMID: 35349301
References
-
- Zhang JM, Wang HK, Ye CQ, Ge W, Chen Y, Jiang ZL, Wu CP, Poo MM, Duan S, ATP released by astrocytes mediates glutamatergic activity-dependent heterosynaptic suppression. Neuron 40, 971–982 (2003). - PubMed
-
- Park KH, Rubin LE, Gross SS, Levi R, Nitric oxide is a mediator of hypoxic coronary vasodilatation. Relation to adenosine and cyclooxygenase-derived metabolites. Circ. Res 71, 992–1001 (1992). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
