

Scaling the leaf length-times-width equation to predict total leaf area of shoots
- PMID: 35350072
- PMCID: PMC9445601
- DOI: 10.1093/aob/mcac043
Scaling the leaf length-times-width equation to predict total leaf area of shoots
Abstract
Background and aims: An individual plant consists of different-sized shoots, each of which consists of different-sized leaves. To predict plant-level physiological responses from the responses of individual leaves, modelling this within-shoot leaf size variation is necessary. Within-plant leaf trait variation has been well investigated in canopy photosynthesis models but less so in plant allometry. Therefore, integration of these two different approaches is needed.
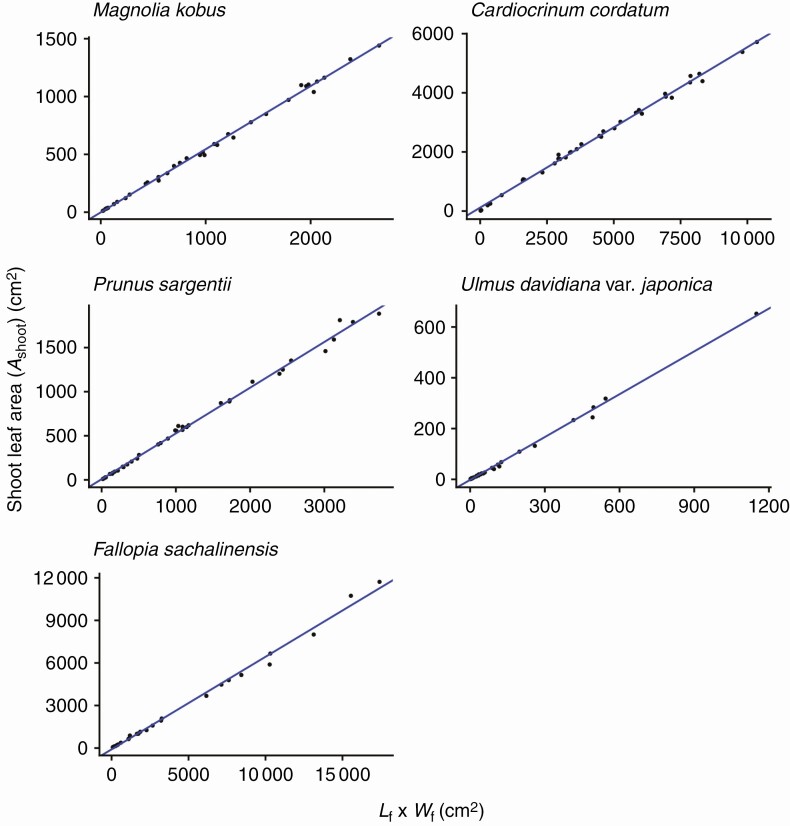
Methods: We focused on an established leaf-level relationship that the area of an individual leaf lamina is proportional to the product of its length and width. The geometric interpretation of this equation is that different-sized leaf laminas from a single species share the same basic form. Based on this shared basic form, we synthesized a new length-times-width equation predicting total shoot leaf area from the collective dimensions of leaves that comprise a shoot. Furthermore, we showed that several previously established empirical relationships, including the allometric relationships between total shoot leaf area, maximum individual leaf length within the shoot and total leaf number of the shoot, can be unified under the same geometric argument. We tested the model predictions using five species, all of which have simple leaves, selected from diverse taxa (Magnoliids, monocots and eudicots) and from different growth forms (trees, erect herbs and rosette herbs).
Key results: For all five species, the length-times-width equation explained within-species variation of total leaf area of a shoot with high accuracy (R2 > 0.994). These strong relationships existed despite leaf dimensions scaling very differently between species. We also found good support for all derived predictions from the model (R2 > 0.85).
Conclusions: Our model can be incorporated to improve previous models of allometry that do not consider within-shoot size variation of individual leaves, providing a cross-scale linkage between individual leaf-size variation and shoot-size variation.
Keywords: Cardiocrinum cordatum; Fallopia sachalinensis; Magnolia kobus; Prunus sargentii; Ulmus davidiana var. japonica; Allometry; Corner’s rule; intraspecific; leaf size; scaling; self-affine; shoot size.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures









References
-
- Anten NPR. 2016. Optimization and game theory in canopy models. In: Hikosaka K, Niinemets Ü, Anten NPR, eds. Canopy photosynthesis: from basics to applications. Dordrecht: Springer Netherlands, 355–377.
-
- APG IV. 2016. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society 181: 1–20.
-
- Auguie B. 2017. https://CRAN.R-project.org/package=gridExtra.
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
