

The In Vivo and In Vitro Architecture of the Hepatitis C Virus RNA Genome Uncovers Functional RNA Secondary and Tertiary Structures
- PMID: 35353000
- PMCID: PMC9044954
- DOI: 10.1128/jvi.01946-21
The In Vivo and In Vitro Architecture of the Hepatitis C Virus RNA Genome Uncovers Functional RNA Secondary and Tertiary Structures
Abstract
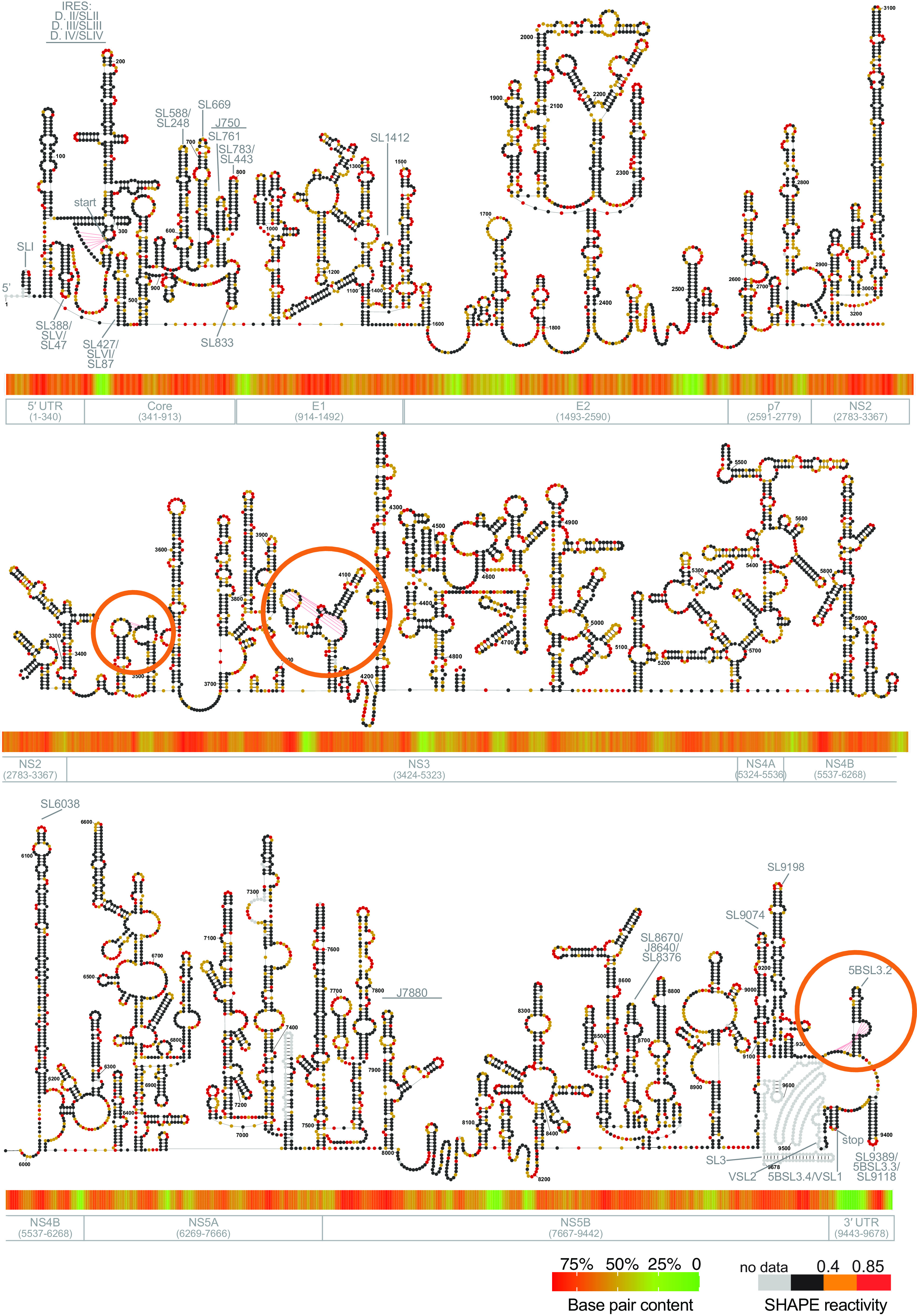
Hepatitis C virus (HCV) is a positive-strand RNA virus that remains one of the main contributors to chronic liver disease worldwide. Studies over the last 30 years have demonstrated that HCV contains a highly structured RNA genome and many of these structures play essential roles in the HCV life cycle. Despite the importance of riboregulation in this virus, most of the HCV RNA genome remains functionally unstudied. Here, we report a complete secondary structure map of the HCV RNA genome in vivo, which was studied in parallel with the secondary structure of the same RNA obtained in vitro. Our results show that HCV is folded extensively in the cellular context. By performing comprehensive structural analyses on both in vivo data and in vitro data, we identify compact and conserved secondary and tertiary structures throughout the genome. Genetic and evolutionary functional analyses demonstrate that many of these elements play important roles in the virus life cycle. In addition to providing a comprehensive map of RNA structures and riboregulatory elements in HCV, this work provides a resource for future studies aimed at identifying therapeutic targets and conducting further mechanistic studies on this important human pathogen. IMPORTANCE HCV has one of the most highly structured RNA genomes studied to date, and it is a valuable model system for studying the role of RNA structure in protein-coding genes. While previous studies have identified individual cases of regulatory RNA structures within the HCV genome, the full-length structure of the HCV genome has not been determined in vivo. Here, we present the complete secondary structure map of HCV determined both in cells and from corresponding transcripts generated in vitro. In addition to providing a comprehensive atlas of functional secondary structural elements throughout the genomic RNA, we identified a novel set of tertiary interactions and demonstrated their functional importance. In terms of broader implications, the pipeline developed in this study can be applied to other long RNAs, such as long noncoding RNAs. In addition, the RNA structural motifs characterized in this study broaden the repertoire of known riboregulatory elements.
Keywords: HCV; RNA folding; SHAPE; pseudoknot; viral replication.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







Similar articles
-
Comprehensive survey of conserved RNA secondary structures in full-genome alignment of Hepatitis C virus.Sci Rep. 2024 Jul 2;14(1):15145. doi: 10.1038/s41598-024-62897-0. Sci Rep. 2024. PMID: 38956134 Free PMC article.
-
The Coding Region of the HCV Genome Contains a Network of Regulatory RNA Structures.Mol Cell. 2016 Apr 7;62(1):111-20. doi: 10.1016/j.molcel.2016.01.024. Epub 2016 Feb 25. Mol Cell. 2016. PMID: 26924328 Free PMC article.
-
Genetic Analysis of Serum-Derived Defective Hepatitis C Virus Genomes Revealed Novel Viral cis Elements for Virus Replication and Assembly.J Virol. 2018 Mar 14;92(7):e02182-17. doi: 10.1128/JVI.02182-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29367245 Free PMC article.
-
Cellular and molecular biology of HCV infection and hepatitis.Clin Sci (Lond). 2009 Jun 15;117(2):49-65. doi: 10.1042/CS20080631. Clin Sci (Lond). 2009. PMID: 19515018 Review.
-
Functional RNA structures throughout the Hepatitis C Virus genome.Curr Opin Virol. 2017 Jun;24:79-86. doi: 10.1016/j.coviro.2017.04.007. Epub 2017 May 13. Curr Opin Virol. 2017. PMID: 28511116 Free PMC article. Review.
Cited by
-
The pseudoknot structure of a viral RNA reveals a conserved mechanism for programmed exoribonuclease resistance.bioRxiv [Preprint]. 2024 Dec 18:2024.12.17.628992. doi: 10.1101/2024.12.17.628992. bioRxiv. 2024. PMID: 39763890 Free PMC article. Preprint.
-
Cell Compartment-Specific Folding of Ty1 Long Terminal Repeat Retrotransposon RNA Genome.Viruses. 2022 Sep 10;14(9):2007. doi: 10.3390/v14092007. Viruses. 2022. PMID: 36146813 Free PMC article.
-
Systematic detection of tertiary structural modules in large RNAs and RNP interfaces by Tb-seq.Nat Commun. 2023 Jun 9;14(1):3426. doi: 10.1038/s41467-023-38623-1. Nat Commun. 2023. PMID: 37296103 Free PMC article.
-
Structural analysis of the lncRNA SChLAP1 reveals protein binding interfaces and a conformationally heterogenous retroviral insertion.RNA. 2025 Aug 18;31(9):1260-1286. doi: 10.1261/rna.080488.125. RNA. 2025. PMID: 40500149 Free PMC article.
-
Micro RNAs as a potential biomarker for predicting liver fibrosis severity in hepatitis C virus affected patients.World J Virol. 2025 Jun 25;14(2):101976. doi: 10.5501/wjv.v14.i2.101976. World J Virol. 2025. PMID: 40575638 Free PMC article.
References
-
- Bartenschlager R. 2013. Hepatitis C virus: from molecular virology to antiviral therapy preface. Curr Top Microbiol Immunol 369:V–Vi.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
