

The two pore potassium channel THIK-1 regulates NLRP3 inflammasome activation
- PMID: 35353387
- PMCID: PMC9314991
- DOI: 10.1002/glia.24174
The two pore potassium channel THIK-1 regulates NLRP3 inflammasome activation
Abstract
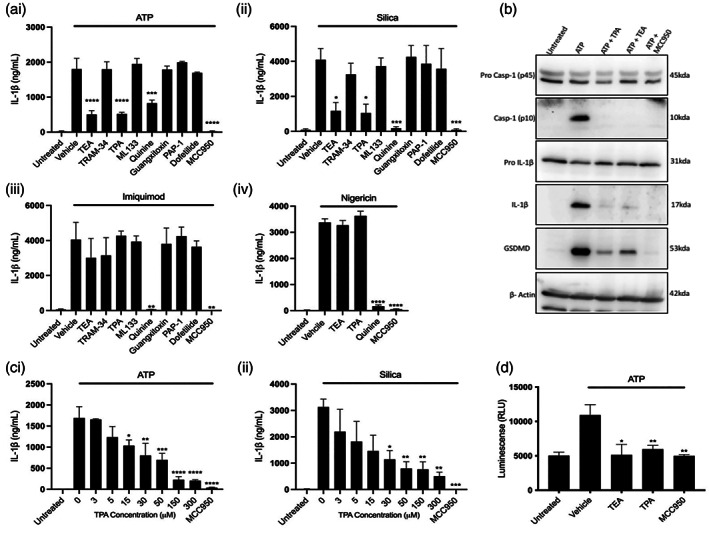
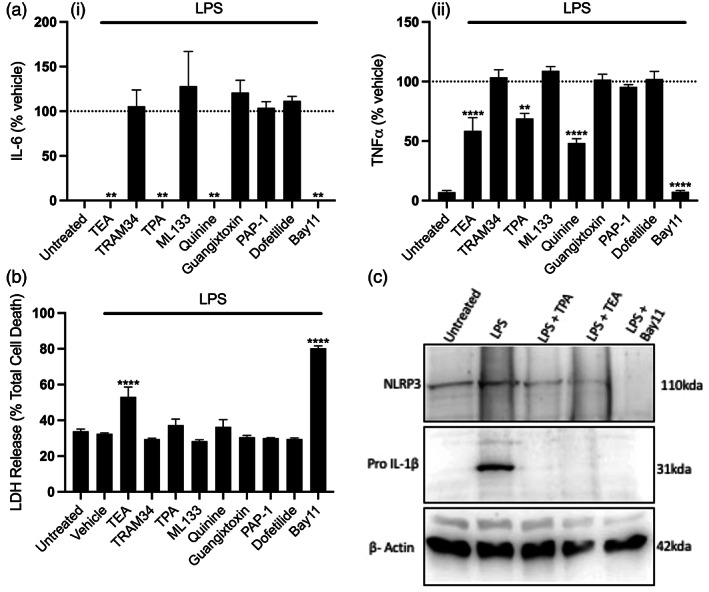
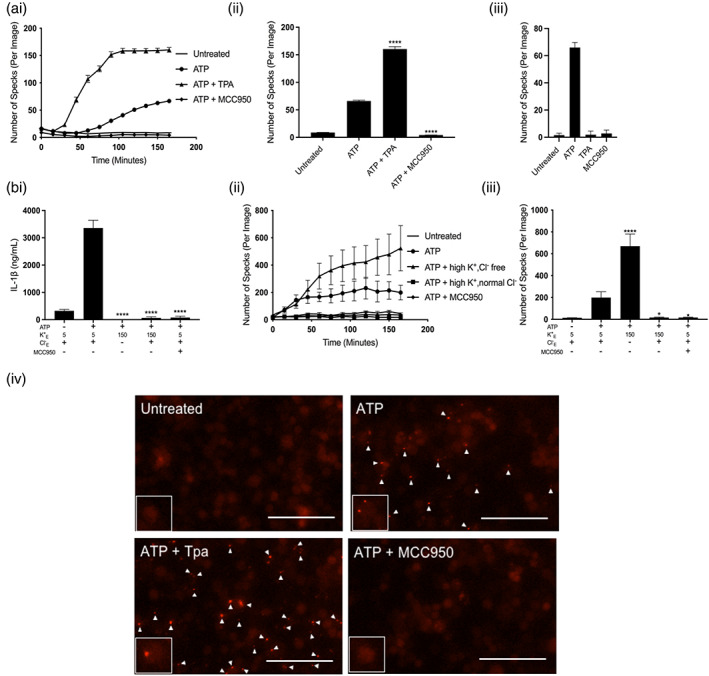
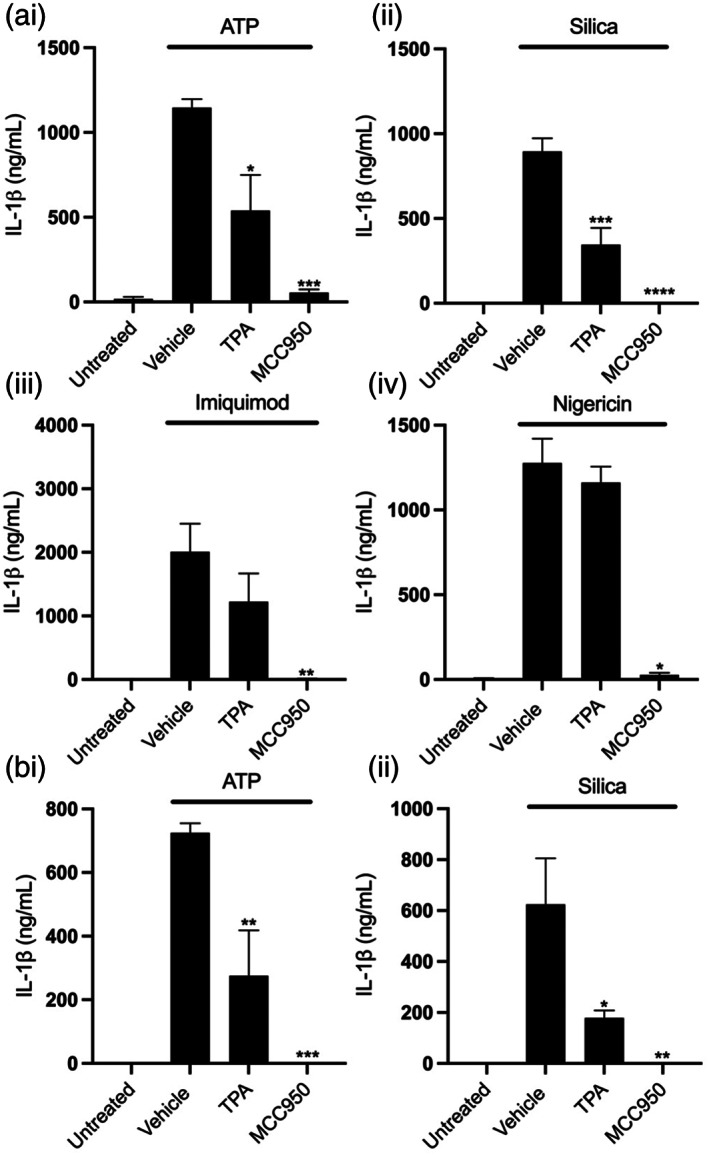
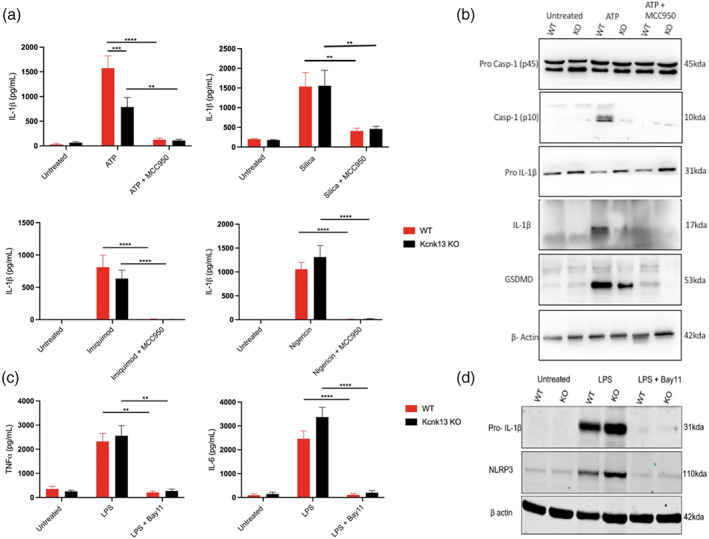
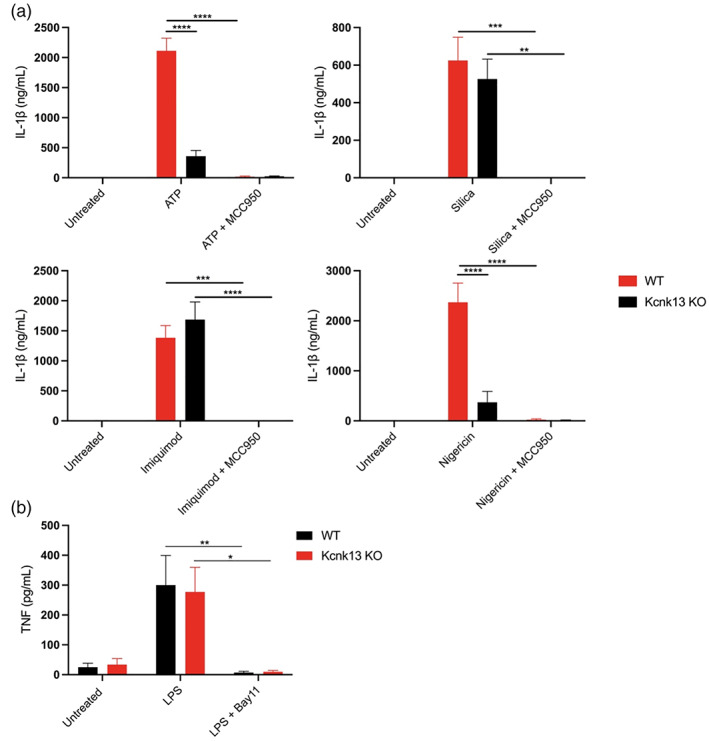
The NLRP3 (NLR family, pyrin domain containing 3) inflammasome is a multi-protein complex responsible for the activation of caspase-1 and the subsequent cleavage and activation of the potent proinflammatory cytokines IL-1β and IL-18, and pyroptotic cell death. NLRP3 is implicated as a driver of inflammation in a range of disorders including neurodegenerative diseases, type 2 diabetes, and atherosclerosis. A commonly reported mechanism contributing to NLRP3 inflammasome activation is potassium ion (K+ ) efflux across the plasma membrane. Identification of K+ channels involved in NLRP3 activation remains incomplete. Here, we investigated the role of the K+ channel THIK-1 in NLRP3 activation. Both pharmacological inhibitors and cells from THIK-1 knockout (KO) mice were used to assess THIK-1 contribution to macrophage NLRP3 activation in vitro. Pharmacological inhibition of THIK-1 inhibited caspase-1 activation and IL-1β release from mouse bone-marrow-derived macrophages (BMDMs), mixed glia, and microglia in response to NLRP3 agonists. Similarly, BMDMs and microglia from THIK-1 KO mice had reduced NLRP3-dependent IL-1β release in response to P2X7 receptor activation with ATP. Overall, these data suggest that THIK-1 is a regulator of NLRP3 inflammasome activation in response to ATP and identify THIK-1 as a potential therapeutic target for inflammatory disease.
Keywords: NLRP3; THIK-1; inflammasome; inflammation; interleukin-1; potassium channel.
© 2022 The Authors. GLIA published by Wiley Periodicals LLC.
Figures
References
-
- Akopova, O. V. (2017). Direct and off‐target effects of ATP‐sensitive potassium channels opener diazoxide. Journal of Drug Metabolism & Toxicology, 8, 227. 10.4172/2157-7609.1000227 - DOI
-
- Baldwin, A. G. , Rivers‐Auty, J. , Daniels, M. J. D. , White, C. S. , Schwalbe, C. H. , Schilling, T. , Hammadi, H. , Jaiyong, P. , Spencer, N. G. , England, H. , Luheshi, N. M. , Kadirvel, M. , Lawrence, C. B. , Rothwell, N. J. , Harte, M. K. , Bryce, R. A. , Allan, S. M. , Eder, C. , Freeman, S. , & Brough, D. (2017). Boron‐based inhibitors of the NLRP3 Inflammasome. Cell Chemical Biology, 24, 1321–1335. 10.1016/j.chembiol.2017.08.011 - DOI - PMC - PubMed
-
- Baron, L. , Gombault, A. , Fanny, M. , Villeret, B. , Savigny, F. , Guillou, N. , Panek, C. , Le Bert, M. , Lagente, V. , Rassendren, F. , Riteau, N. , & Couillin, I. (2015). The NLRP3 inflammasome is activated by nanoparticles through ATP, ADP and adenosine. Cell Death and Disease, 6, e1629. 10.1038/cddis.2014.576 - DOI - PMC - PubMed
-
- Bauernfeind, F. , Horvath, G. , Stutz, A. , Alnemri, E. S. , Speert, D. , Fernandes‐alnemri, T. , Wu, J. , Brian, G. , Fitzgerald, K. A. , Hornung, V. , & Latz, E. (2009). NF‐kB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. Journal of Immunology, 183, 787–791. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
