

Complete genomic and epigenetic maps of human centromeres
- PMID: 35357911
- PMCID: PMC9233505
- DOI: 10.1126/science.abl4178
Complete genomic and epigenetic maps of human centromeres
Abstract
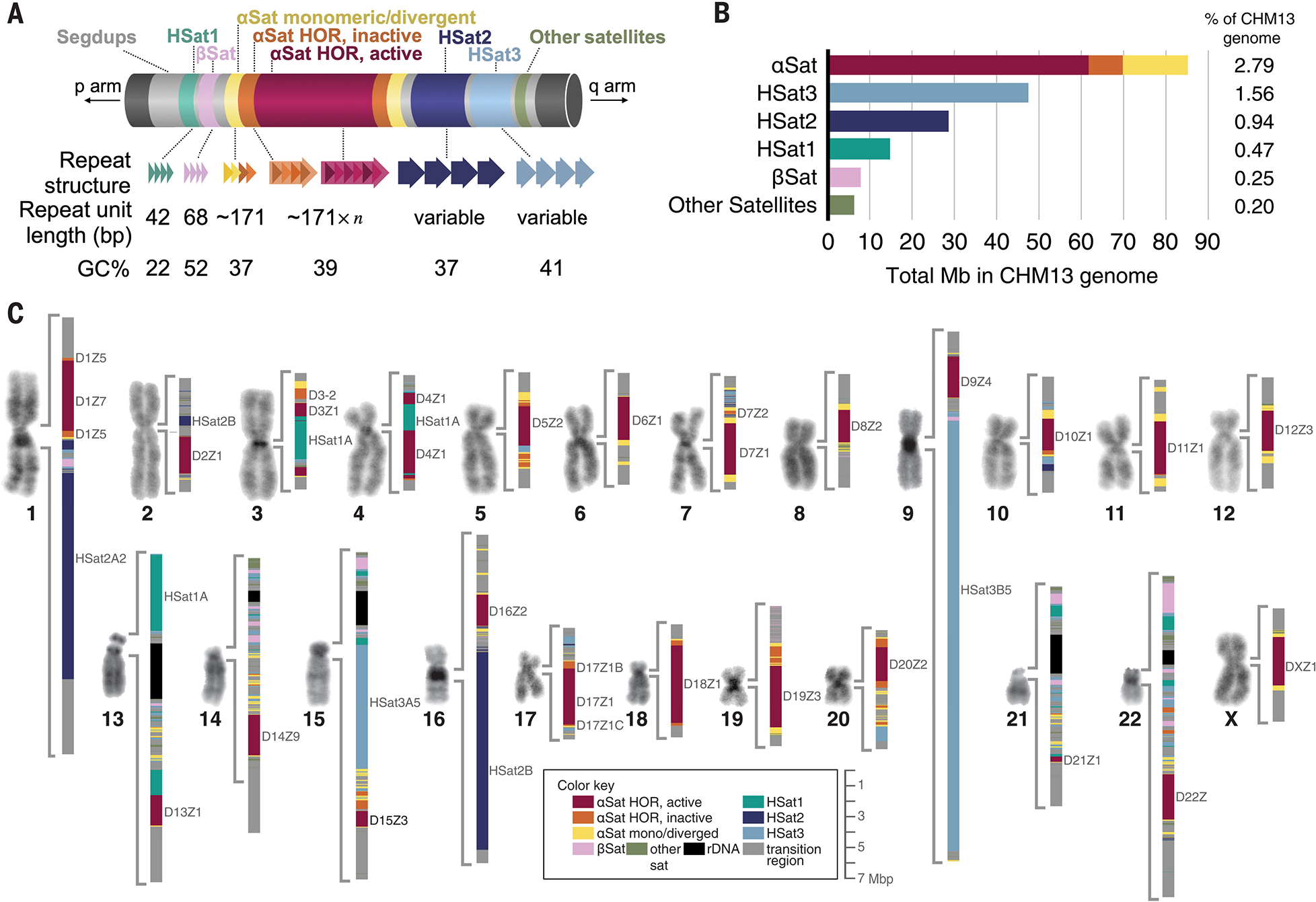
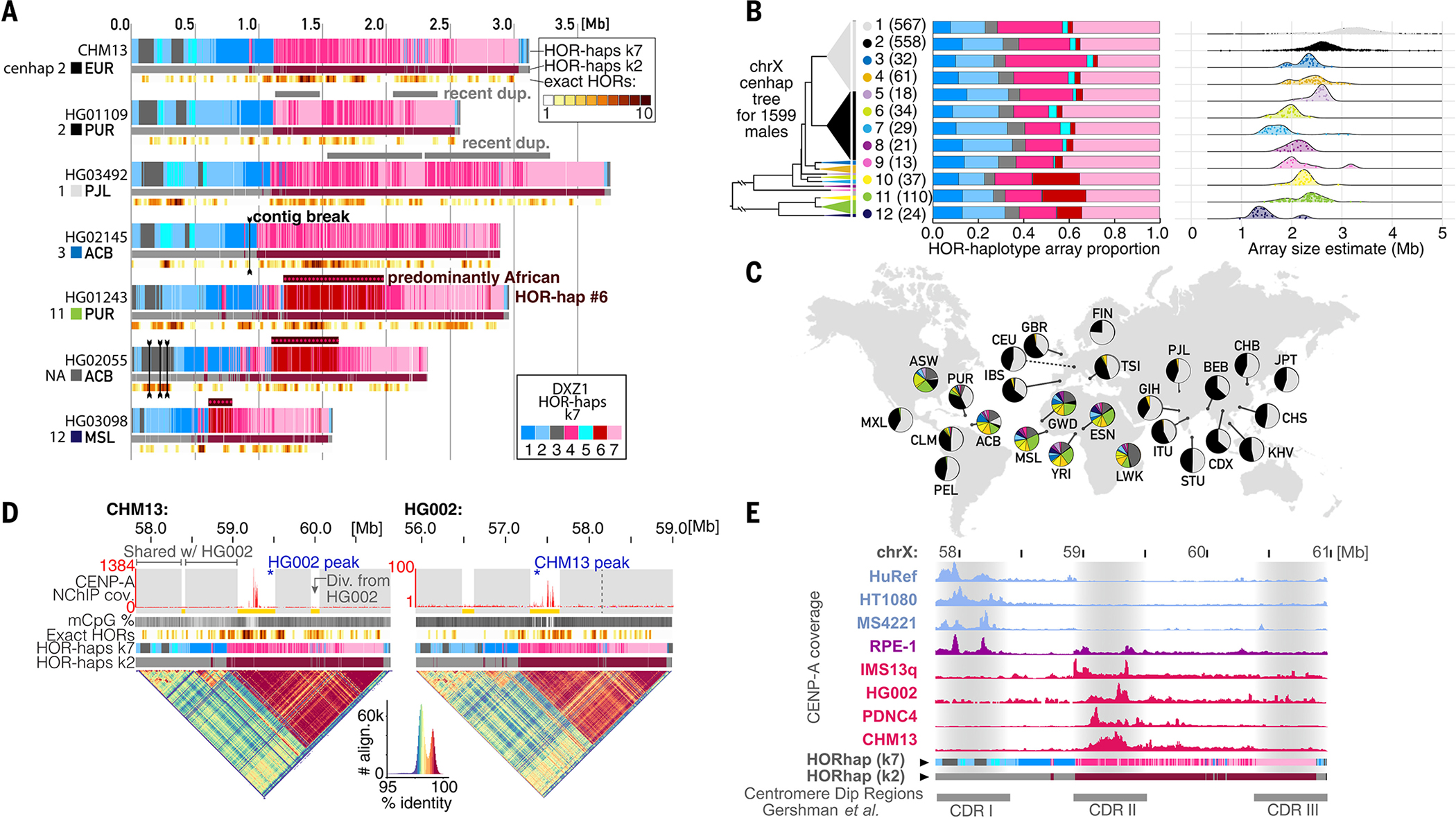
Existing human genome assemblies have almost entirely excluded repetitive sequences within and near centromeres, limiting our understanding of their organization, evolution, and functions, which include facilitating proper chromosome segregation. Now, a complete, telomere-to-telomere human genome assembly (T2T-CHM13) has enabled us to comprehensively characterize pericentromeric and centromeric repeats, which constitute 6.2% of the genome (189.9 megabases). Detailed maps of these regions revealed multimegabase structural rearrangements, including in active centromeric repeat arrays. Analysis of centromere-associated sequences uncovered a strong relationship between the position of the centromere and the evolution of the surrounding DNA through layered repeat expansions. Furthermore, comparisons of chromosome X centromeres across a diverse panel of individuals illuminated high degrees of structural, epigenetic, and sequence variation in these complex and rapidly evolving regions.
Conflict of interest statement
Figures



Comment in
-
The final pieces of the human genome.Nat Rev Genet. 2022 Jun;23(6):321. doi: 10.1038/s41576-022-00494-5. Nat Rev Genet. 2022. PMID: 35488041 No abstract available.
References
Publication types
MeSH terms
Grants and funding
- U24 HG010263/HG/NHGRI NIH HHS/United States
- U01 HG010961/HG/NHGRI NIH HHS/United States
- DP2 MH119424/MH/NIMH NIH HHS/United States
- R35 GM139653/GM/NIGMS NIH HHS/United States
- R21 CA240199/CA/NCI NIH HHS/United States
- OT2 OD026682/OD/NIH HHS/United States
- R01 GM074728/GM/NIGMS NIH HHS/United States
- R21 CA238758/CA/NCI NIH HHS/United States
- U01 HG010971/HG/NHGRI NIH HHS/United States
- P20 GM103546/GM/NIGMS NIH HHS/United States
- R01 GM132600/GM/NIGMS NIH HHS/United States
- R01 GM124041/GM/NIGMS NIH HHS/United States
- T32 GM007445/GM/NIGMS NIH HHS/United States
- U41 HG010972/HG/NHGRI NIH HHS/United States
- F31 HG011205/HG/NHGRI NIH HHS/United States
- F32 GM134558/GM/NIGMS NIH HHS/United States
- R01 HG010169/HG/NHGRI NIH HHS/United States
- R01 GM129263/GM/NIGMS NIH HHS/United States
- U41 HG007234/HG/NHGRI NIH HHS/United States
- U01 CA253481/CA/NCI NIH HHS/United States
- U24 HG007234/HG/NHGRI NIH HHS/United States
- R01 HG011274/HG/NHGRI NIH HHS/United States
- U24 HG006620/HG/NHGRI NIH HHS/United States
- R01 HG002939/HG/NHGRI NIH HHS/United States
- R01 AG054712/AG/NIA NIH HHS/United States
- U24 HG010136/HG/NHGRI NIH HHS/United States
- R01 HG010329/HG/NHGRI NIH HHS/United States
- R21 HG010548/HG/NHGRI NIH HHS/United States
- U24 HG011853/HG/NHGRI NIH HHS/United States
- R01 GM123312/GM/NIGMS NIH HHS/United States
- R01 HG009190/HG/NHGRI NIH HHS/United States
- R01 GM117420/GM/NIGMS NIH HHS/United States
- R01 HG010485/HG/NHGRI NIH HHS/United States
- R24 DK106766/DK/NIDDK NIH HHS/United States
