

Effects of Receptor Specificity and Conformational Stability of Influenza A Virus Hemagglutinin on Infection and Activation of Different Cell Types in Human PBMCs
- PMID: 35359920
- PMCID: PMC8963867
- DOI: 10.3389/fimmu.2022.827760
Effects of Receptor Specificity and Conformational Stability of Influenza A Virus Hemagglutinin on Infection and Activation of Different Cell Types in Human PBMCs
Abstract
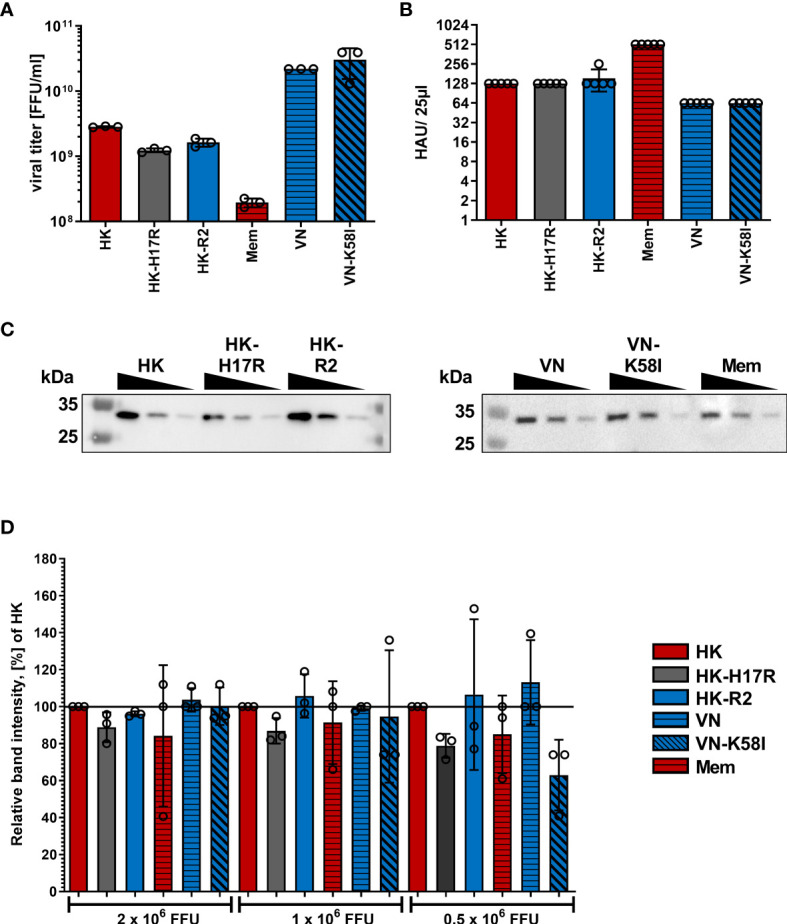
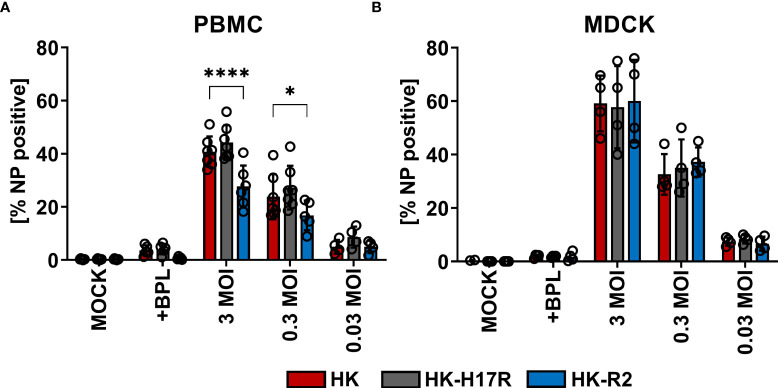
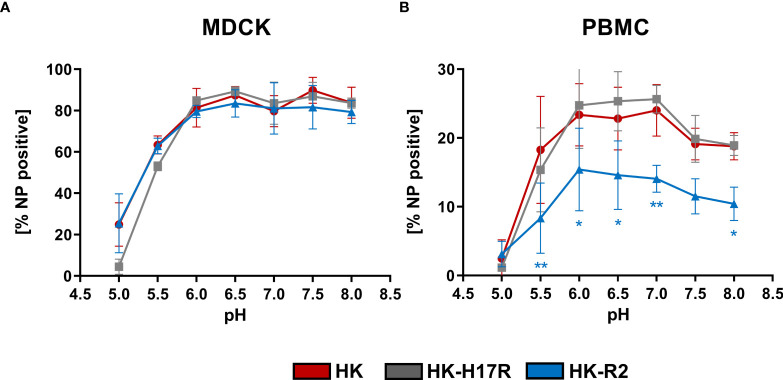
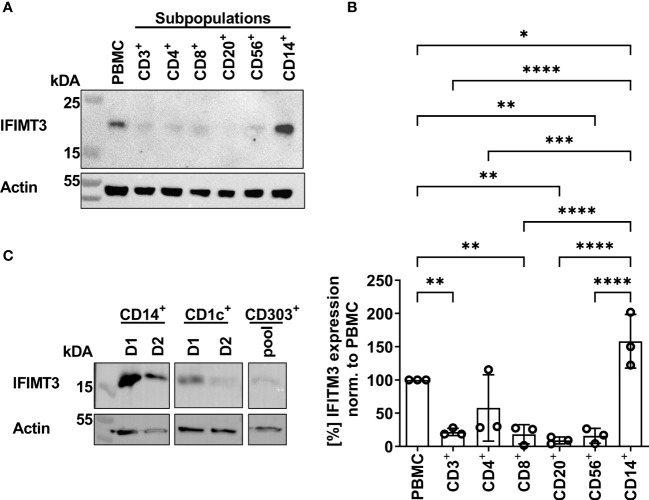
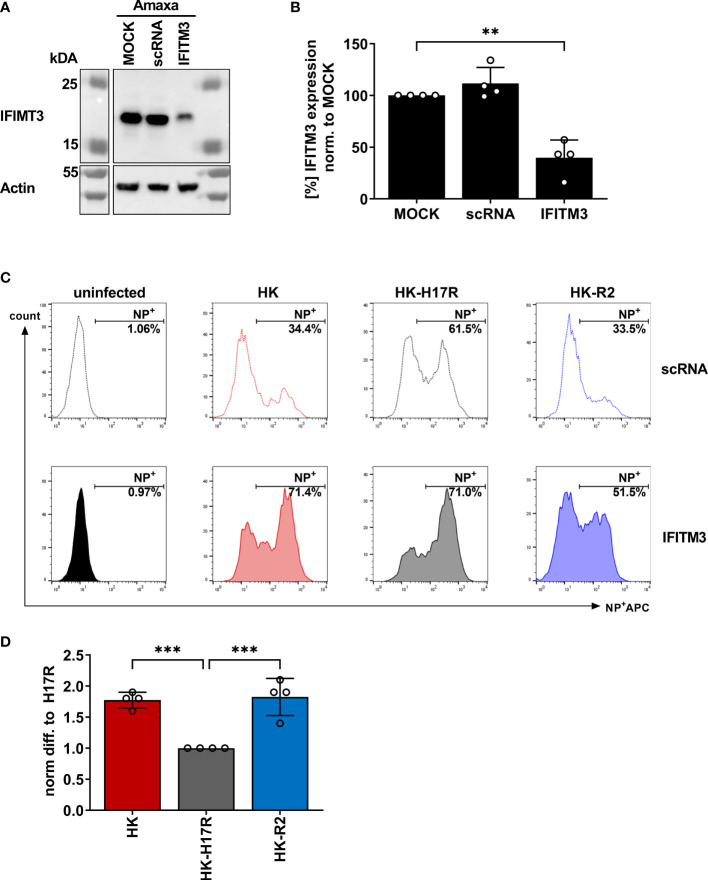
Humans can be infected by zoonotic avian, pandemic and seasonal influenza A viruses (IAVs), which differ by receptor specificity and conformational stability of their envelope glycoprotein hemagglutinin (HA). It was shown that receptor specificity of the HA determines the tropism of IAVs to human airway epithelial cells, the primary target of IAVs in humans. Less is known about potential effects of the HA properties on viral attachment, infection and activation of human immune cells. To address this question, we studied the infection of total human peripheral blood mononuclear cells (PBMCs) and subpopulations of human PBMCs with well characterized recombinant IAVs differing by the HA and the neuraminidase (NA) but sharing all other viral proteins. Monocytes and all subpopulations of lymphocytes were significantly less susceptible to infection by IAVs with avian-like receptor specificity as compared to human-like IAVs, whereas plasmacytoid dendritic cells (pDCs) and myeloid dendritic cells were equally susceptible to IAVs with avian-like and human-like receptor specificity. This tropism correlated with the surface expression of 2-3-linked sialic acids (avian-type receptors) and 2-6-linked sialic acids (human-type receptors). Despite a reduced infectivity of avian-like IAVs for PBMCs, these viruses were not less efficient than human-like IAVs in terms of cell activation as judged by the induction of cellular mRNA of IFN-α, CCL5, RIG-I, and IL-6. Elevated levels of IFN-α mRNA were accompanied by elevated IFN-α protein secretion in primary human pDC. We found that high basal expression in monocytes of antiviral interferon-induced transmembrane protein 3 (IFITM3) limited viral infection in these cells. siRNA-mediated knockdown of IFITM3 in monocytes demonstrated that viral sensitivity to inhibition by IFITM3 correlated with the conformational stability of the HA. Our study provides new insights into the role of host- and strain-specific differences of HA in the interaction of IAVs with human immune cells and advances current understanding of the mechanisms of viral cell tropism, pathogenesis and markers of virulence.
Keywords: hemagglutinin; influenza; primary immune cells; receptor specificity; tropism.
Copyright © 2022 Dorna, Kaufmann, Bockmann, Raifer, West, Matrosovich and Bauer.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
HA-Dependent Tropism of H5N1 and H7N9 Influenza Viruses to Human Endothelial Cells Is Determined by Reduced Stability of the HA, Which Allows the Virus To Cope with Inefficient Endosomal Acidification and Constitutively Expressed IFITM3.J Virol. 2019 Dec 12;94(1):e01223-19. doi: 10.1128/JVI.01223-19. Print 2019 Dec 12. J Virol. 2019. PMID: 31597765 Free PMC article.
-
pH Optimum of Hemagglutinin-Mediated Membrane Fusion Determines Sensitivity of Influenza A Viruses to the Interferon-Induced Antiviral State and IFITMs.J Virol. 2017 May 12;91(11):e00246-17. doi: 10.1128/JVI.00246-17. Print 2017 Jun 1. J Virol. 2017. PMID: 28356532 Free PMC article.
-
Mutation of the second sialic acid-binding site of influenza A virus neuraminidase drives compensatory mutations in hemagglutinin.PLoS Pathog. 2020 Aug 27;16(8):e1008816. doi: 10.1371/journal.ppat.1008816. eCollection 2020 Aug. PLoS Pathog. 2020. PMID: 32853241 Free PMC article.
-
Adaptation of influenza viruses to human airway receptors.J Biol Chem. 2021 Jan-Jun;296:100017. doi: 10.1074/jbc.REV120.013309. Epub 2020 Nov 22. J Biol Chem. 2021. PMID: 33144323 Free PMC article. Review.
-
Receptor binding and pH stability - how influenza A virus hemagglutinin affects host-specific virus infection.Biochim Biophys Acta. 2014 Apr;1838(4):1153-68. doi: 10.1016/j.bbamem.2013.10.004. Epub 2013 Oct 24. Biochim Biophys Acta. 2014. PMID: 24161712 Review.
Cited by
-
STING regulates NETs formation by activating GSDMD in influenza viral pneumonia.Front Immunol. 2025 Jul 1;16:1598902. doi: 10.3389/fimmu.2025.1598902. eCollection 2025. Front Immunol. 2025. PMID: 40666504 Free PMC article.
-
From HIV to COVID-19, Molecular mechanisms of pathogens' trade-off and persistence in the community, potential targets for new drug development.Bull Natl Res Cent. 2022;46(1):194. doi: 10.1186/s42269-022-00879-w. Epub 2022 Jul 6. Bull Natl Res Cent. 2022. PMID: 35818410 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources