

DNA methylation signature associated with Bohring-Opitz syndrome: a new tool for functional classification of variants in ASXL genes
- PMID: 35361921
- PMCID: PMC9177544
- DOI: 10.1038/s41431-022-01083-0
DNA methylation signature associated with Bohring-Opitz syndrome: a new tool for functional classification of variants in ASXL genes
Abstract
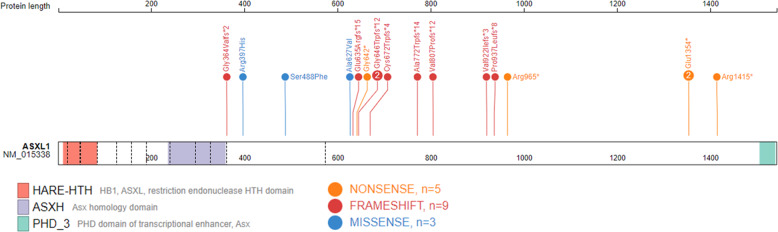
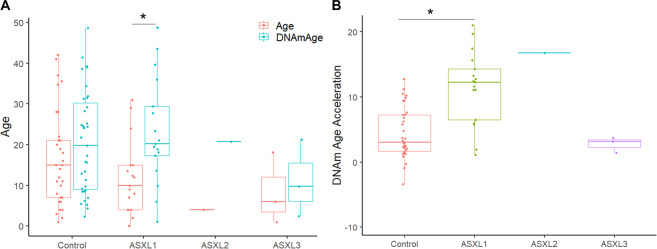
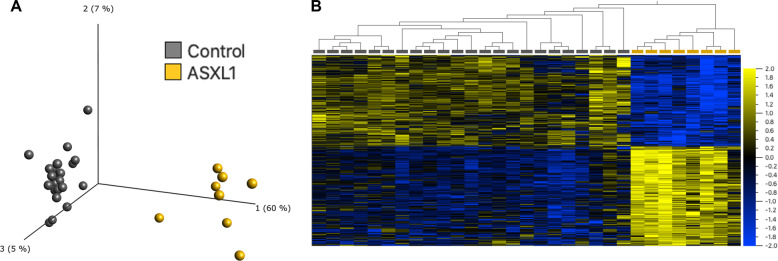
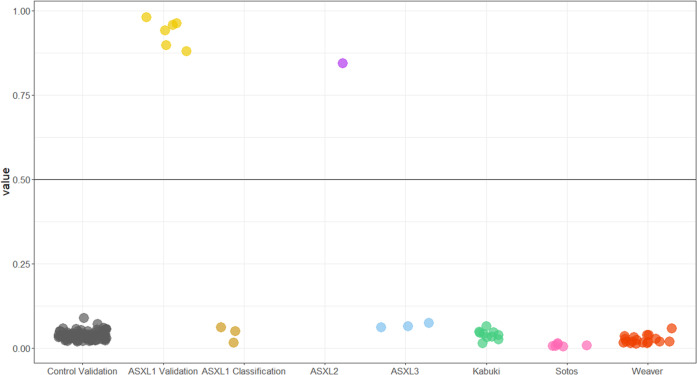
The additional sex combs-like (ASXL) gene family-encoded by ASXL1, ASXL2, and ASXL3-is crucial for mammalian development. Pathogenic variants in the ASXL gene family are associated with three phenotypically distinct neurodevelopmental syndromes. Our previous work has shown that syndromic conditions caused by pathogenic variants in epigenetic regulatory genes show consistent patterns of genome-wide DNA methylation (DNAm) alterations, i.e., DNAm signatures in peripheral blood. Given the role of ASXL1 in chromatin modification, we hypothesized that pathogenic ASXL1 variants underlying Bohring-Opitz syndrome (BOS) have a unique DNAm signature. We profiled whole-blood DNAm for 17 ASXL1 variants, and 35 sex- and age-matched typically developing individuals, using Illumina's Infinium EPIC array. We identified 763 differentially methylated CpG sites in individuals with BOS. Differentially methylated sites overlapped 323 unique genes, including HOXA5 and HOXB4, supporting the functional relevance of DNAm signatures. We used a machine-learning classification model based on the BOS DNAm signature to classify variants of uncertain significance in ASXL1, as well as pathogenic ASXL2 and ASXL3 variants. The DNAm profile of one individual with the ASXL2 variant was BOS-like, whereas the DNAm profiles of three individuals with ASXL3 variants were control-like. We also used Horvath's epigenetic clock, which showed acceleration in DNAm age in individuals with pathogenic ASXL1 variants, and the individual with the pathogenic ASXL2 variant, but not in individuals with ASXL3 variants. These studies enhance our understanding of the epigenetic dysregulation underpinning ASXL gene family-associated syndromes.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
ANKRD11 pathogenic variants and 16q24.3 microdeletions share an altered DNA methylation signature in patients with KBG syndrome.Hum Mol Genet. 2023 Apr 20;32(9):1429-1438. doi: 10.1093/hmg/ddac289. Hum Mol Genet. 2023. PMID: 36440975 Free PMC article.
-
New insights into DNA methylation signatures: SMARCA2 variants in Nicolaides-Baraitser syndrome.BMC Med Genomics. 2019 Jul 9;12(1):105. doi: 10.1186/s12920-019-0555-y. BMC Med Genomics. 2019. PMID: 31288860 Free PMC article.
-
Understanding the phenotypic spectrum of ASXL-related disease: Ten cases and a review of the literature.Am J Med Genet A. 2021 Jun;185(6):1700-1711. doi: 10.1002/ajmg.a.62156. Epub 2021 Mar 10. Am J Med Genet A. 2021. PMID: 33751773 Free PMC article.
-
Functional and cancer genomics of ASXL family members.Br J Cancer. 2013 Jul 23;109(2):299-306. doi: 10.1038/bjc.2013.281. Epub 2013 Jun 4. Br J Cancer. 2013. PMID: 23736028 Free PMC article. Review.
-
Pathogenic ASXL1 somatic variants in reference databases complicate germline variant interpretation for Bohring-Opitz Syndrome.Hum Mutat. 2017 May;38(5):517-523. doi: 10.1002/humu.23203. Epub 2017 Mar 21. Hum Mutat. 2017. PMID: 28229513 Free PMC article. Review.
Cited by
-
Multiomics of Bohring-Opitz syndrome truncating ASXL1 mutations identify canonical and noncanonical Wnt signaling dysregulation.JCI Insight. 2023 May 22;8(10):e167744. doi: 10.1172/jci.insight.167744. JCI Insight. 2023. PMID: 37053013 Free PMC article.
-
KAT6A mutations in Arboleda-Tham syndrome drive epigenetic regulation of posterior HOXC cluster.Hum Genet. 2023 Dec;142(12):1705-1720. doi: 10.1007/s00439-023-02608-3. Epub 2023 Oct 20. Hum Genet. 2023. PMID: 37861717 Free PMC article.
-
ASXL1 truncating variants in BOS and myeloid leukemia drive shared disruption of Wnt-signaling pathways but have differential isoform usage of RUNX3.BMC Med Genomics. 2024 Nov 29;17(1):282. doi: 10.1186/s12920-024-02039-7. BMC Med Genomics. 2024. PMID: 39614348 Free PMC article.
-
Polycomb-associated and Trithorax-associated developmental conditions-phenotypic convergence and heterogeneity.Eur J Hum Genet. 2025 Jan 22. doi: 10.1038/s41431-025-01784-2. Online ahead of print. Eur J Hum Genet. 2025. PMID: 39843918
-
DNA methylation signatures for chromatinopathies: current challenges and future applications.Hum Genet. 2024 Apr;143(4):551-557. doi: 10.1007/s00439-023-02544-2. Epub 2023 Apr 6. Hum Genet. 2024. PMID: 37022461 Free PMC article.
References
-
- Russell BTW, Graham JM Jr. Bohring-Opitz Syndrome. GeneReviews® [Internet] Seattle (WA): University of Washington, Seattle; 1993–2021. - PubMed
