

Crystal structure of the putative cell-wall lipoglycan biosynthesis protein LmcA from Mycobacterium smegmatis
- PMID: 35362472
- PMCID: PMC8972800
- DOI: 10.1107/S2059798322001772
Crystal structure of the putative cell-wall lipoglycan biosynthesis protein LmcA from Mycobacterium smegmatis
Abstract
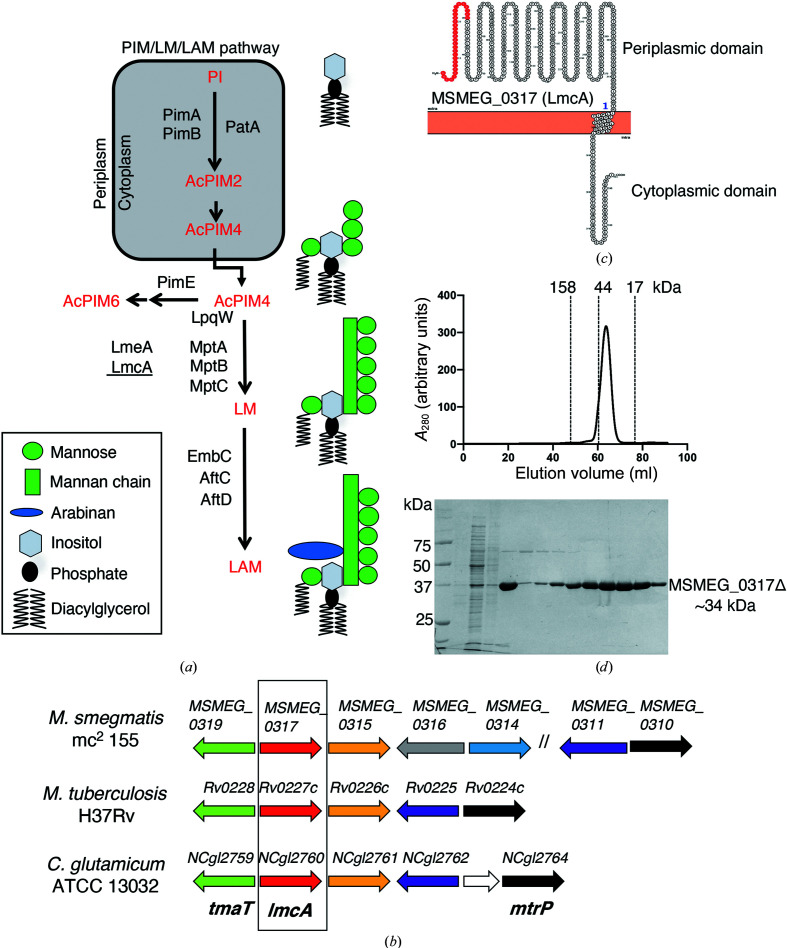
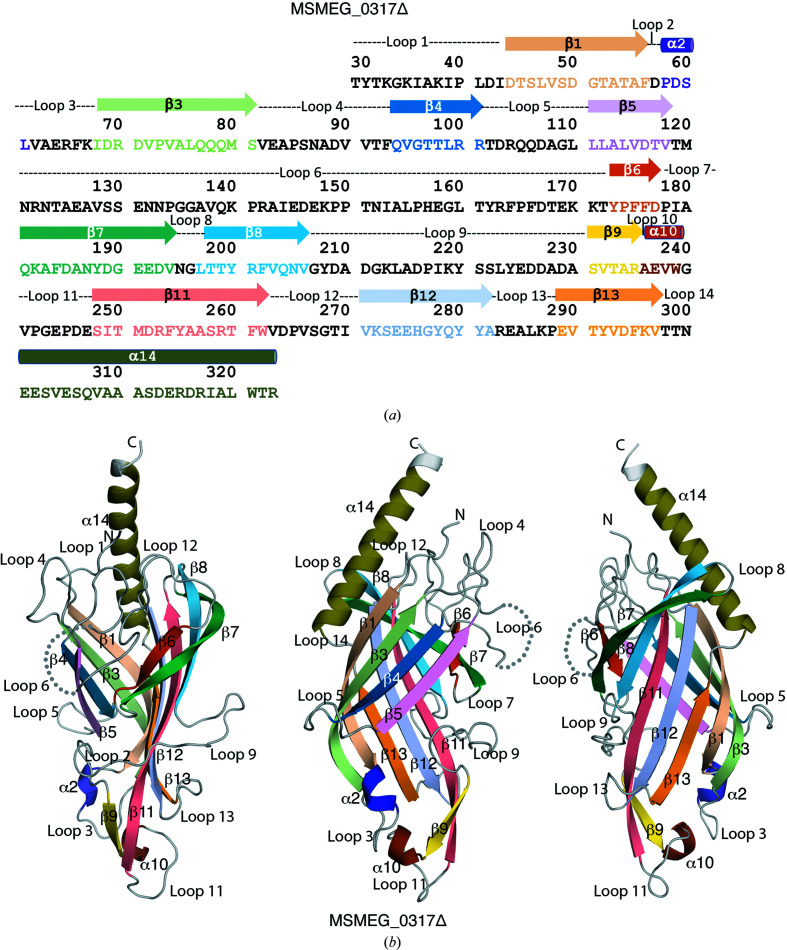
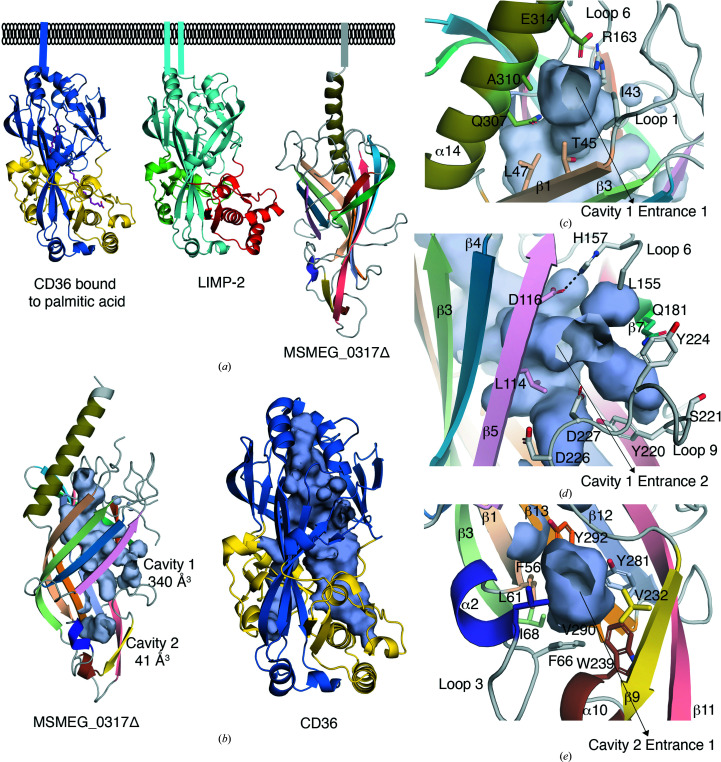
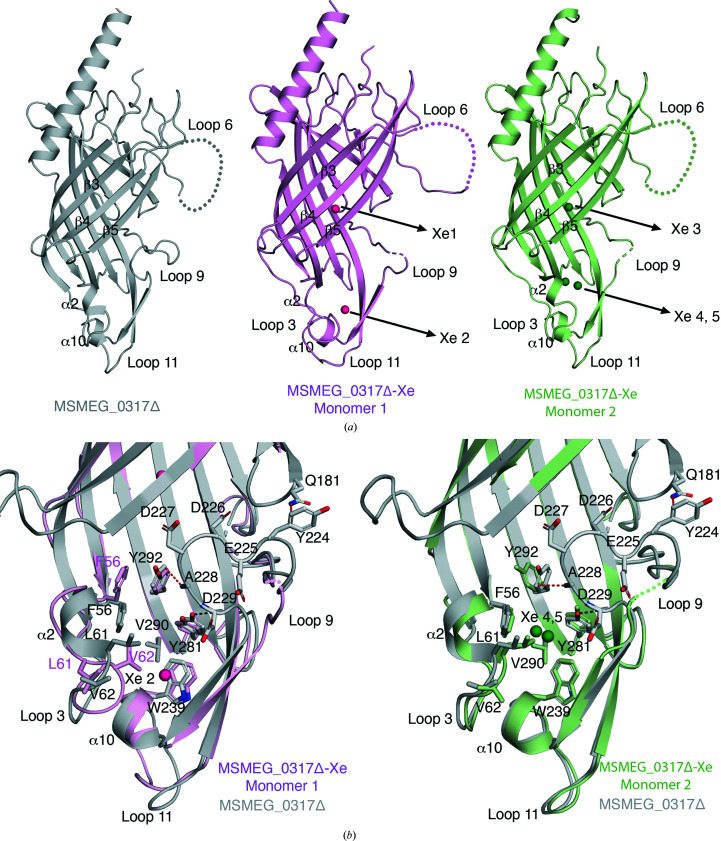
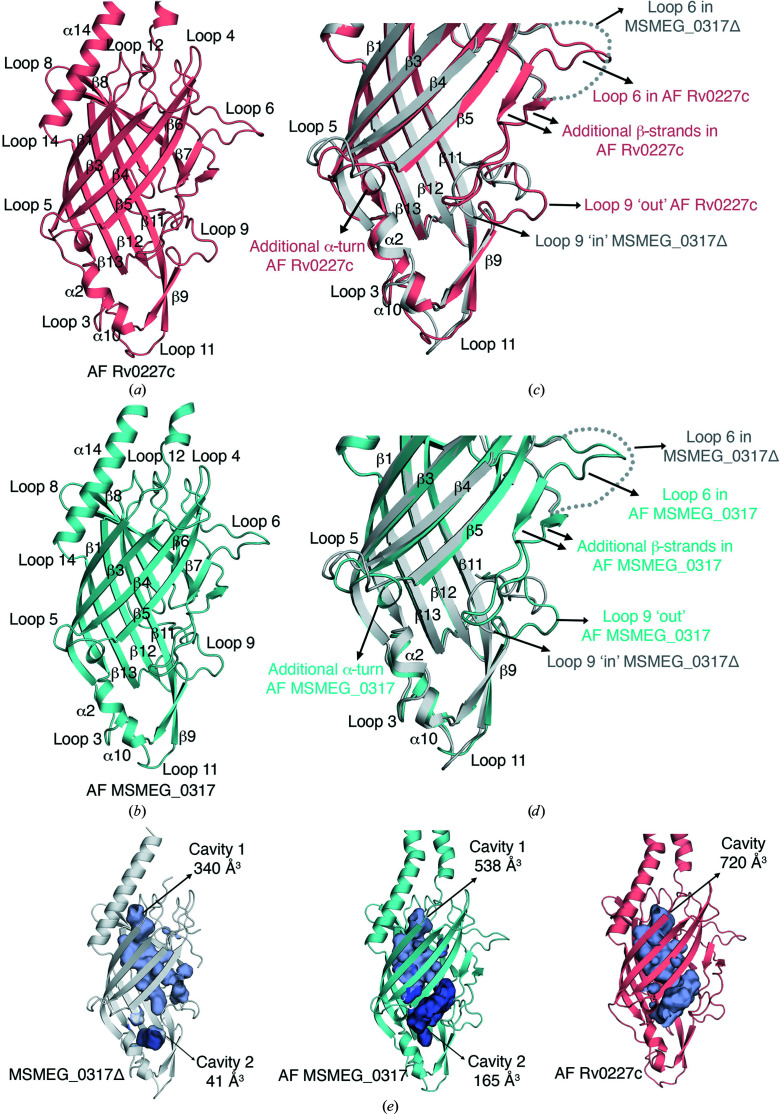
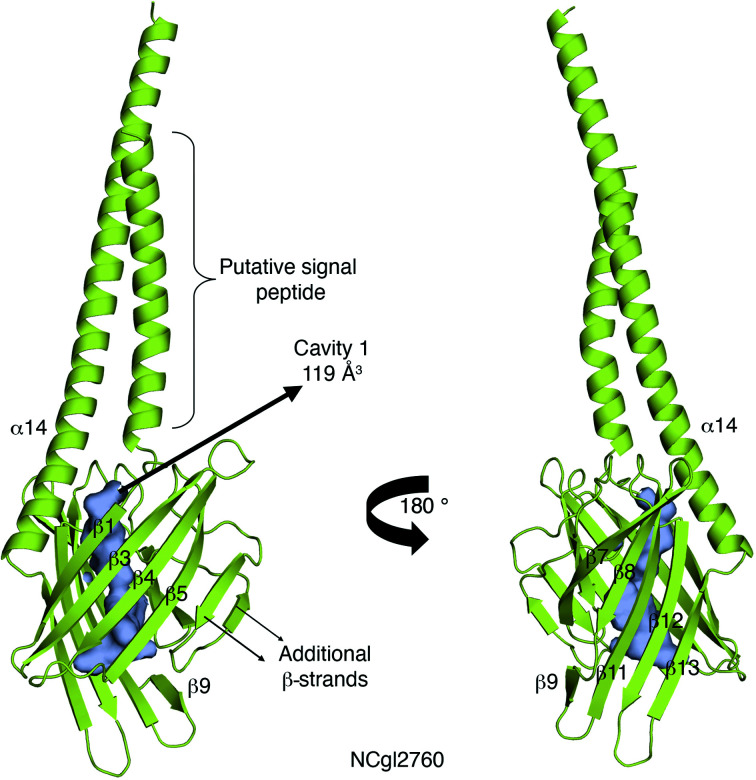
The bacterial genus Mycobacterium includes important pathogens, most notably M. tuberculosis, which infects one-quarter of the entire human population, resulting in around 1.4 million deaths from tuberculosis each year. Mycobacteria, and the closely related corynebacteria, synthesize a class of abundant glycolipids, the phosphatidyl-myo-inositol mannosides (PIMs). PIMs serve as membrane anchors for hyperglycosylated species, lipomannan (LM) and lipoarabinomannan (LAM), which are surface-exposed and modulate the host immune response. Previously, in studies using the model species Corynebacterium glutamicum, NCgl2760 was identified as a novel membrane protein that is required for the synthesis of full-length LM and LAM. Here, the first crystal structure of its ortholog in Mycobacterium smegmatis, MSMEG_0317, is reported at 1.8 Å resolution. The structure revealed an elongated β-barrel fold enclosing two distinct cavities and one α-helix extending away from the β-barrel core, resembling a `cone with a flake' arrangement. Through xenon derivatization and structural comparison with AlphaFold2-derived predictions of the M. tuberculosis homolog Rv0227c, structural elements were identified that may undergo conformational changes to switch from `closed' to `open' conformations, allowing cavity access. An AlphaFold2-derived NCgl2760 model predicted a smaller β-barrel core with an enclosed central cavity, suggesting that all three proteins, which were collectively termed LmcA, may have a common mechanism of ligand binding through these cavities. These findings provide new structural insights into the biosynthetic pathway for a family of surface lipoglycans with important roles in mycobacterial pathogenesis.
Keywords: MSMEG_0317; Mycobacterium smegmatis; Mycobacterium tuberculosis; lipoarabinomannan; lipomannan.
open access.
Figures
References
-
- Brennan, P. J. & Nikaido, H. (1995). Annu. Rev. Biochem. 64, 29–63. - PubMed
-
- Bricogne, G., Blanc, E., Brandl, M., Flensburg, C., Keller, P., Paciorek, W., Roversi, P., Sharff, A., Smart, O. S., Vonrhein, C. & Womack, T. O. (2017). BUSTER. Global Phasing Ltd, Cambridge, United Kingdom.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
