

Coordinated regulation of RNA polymerase II pausing and elongation progression by PAF1
- PMID: 35363521
- PMCID: PMC11093130
- DOI: 10.1126/sciadv.abm5504
Coordinated regulation of RNA polymerase II pausing and elongation progression by PAF1
Abstract
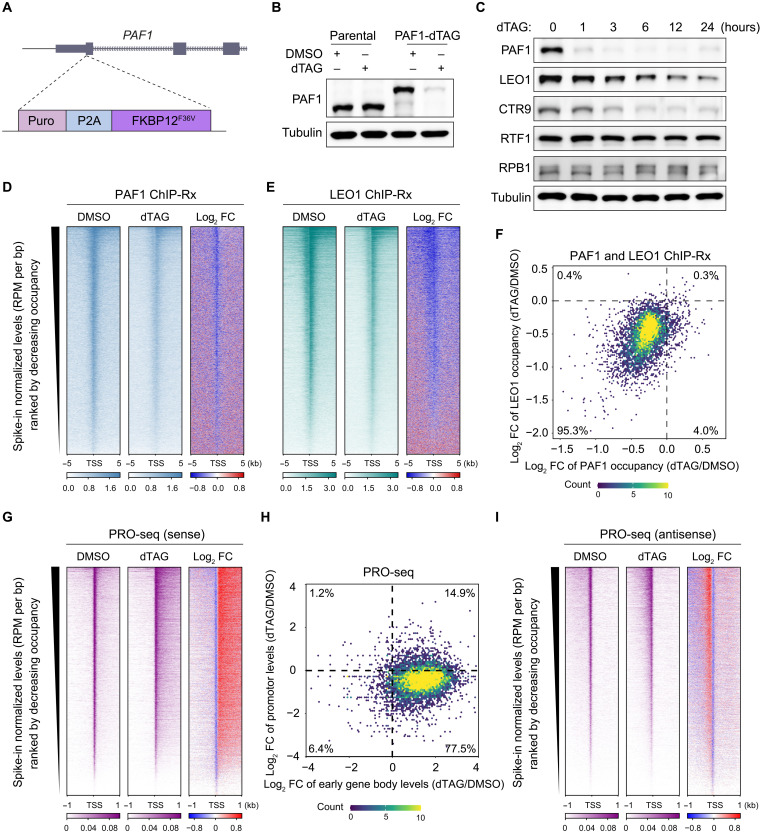
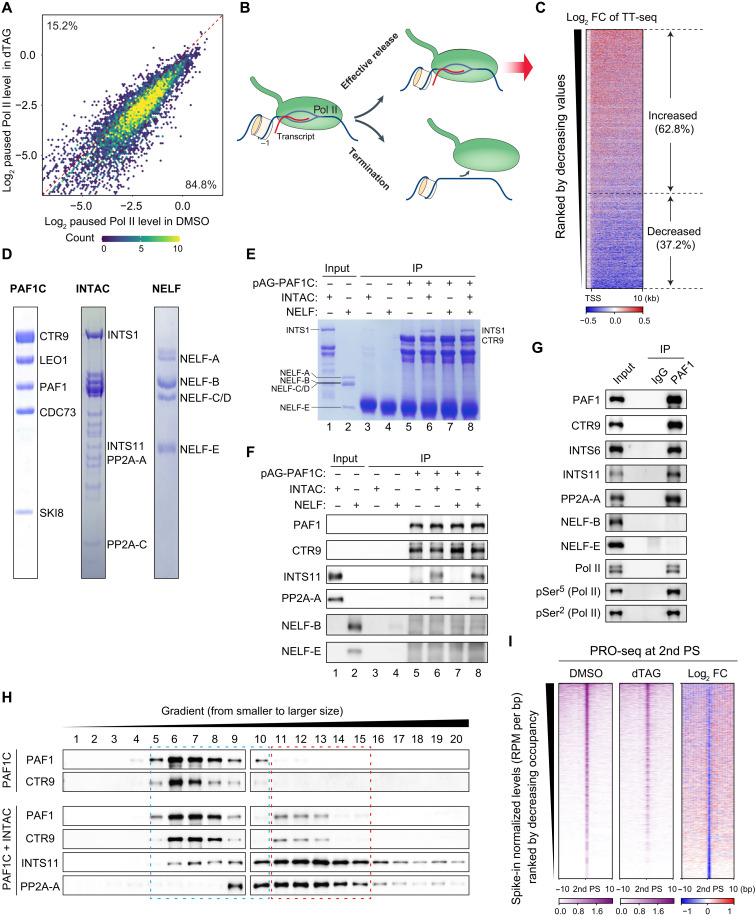
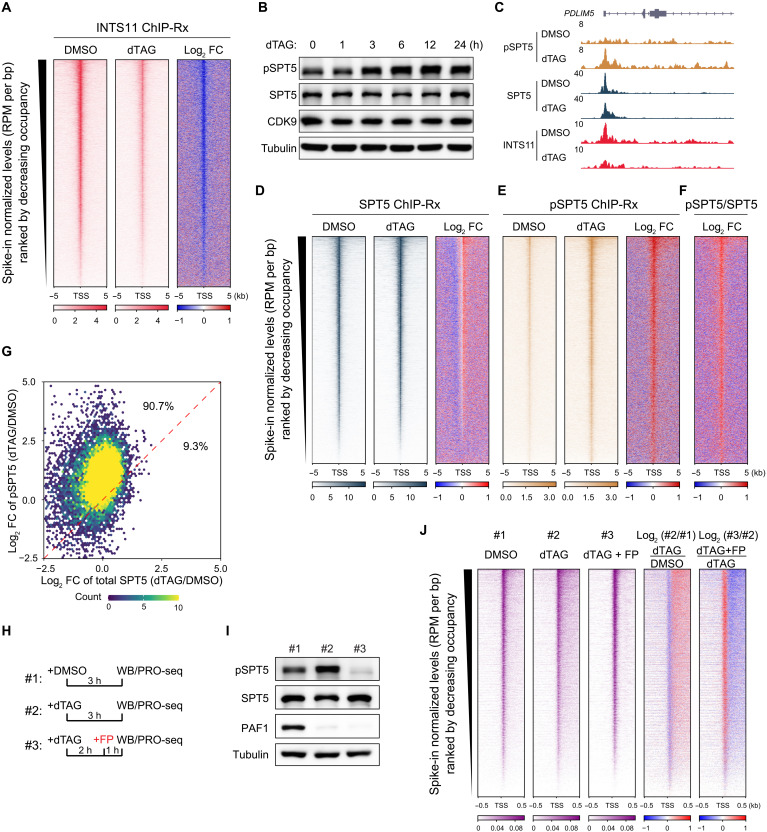
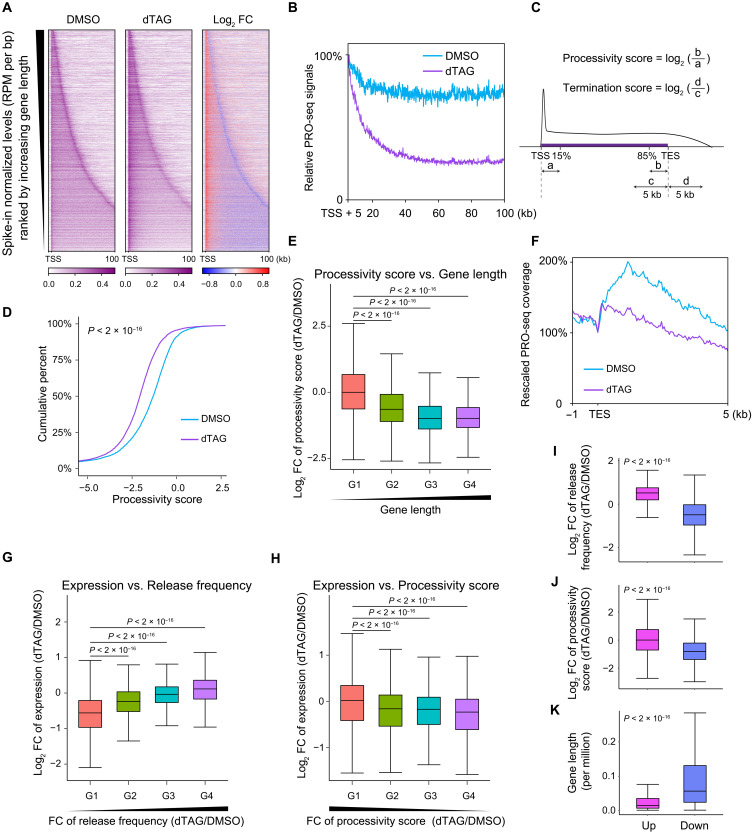
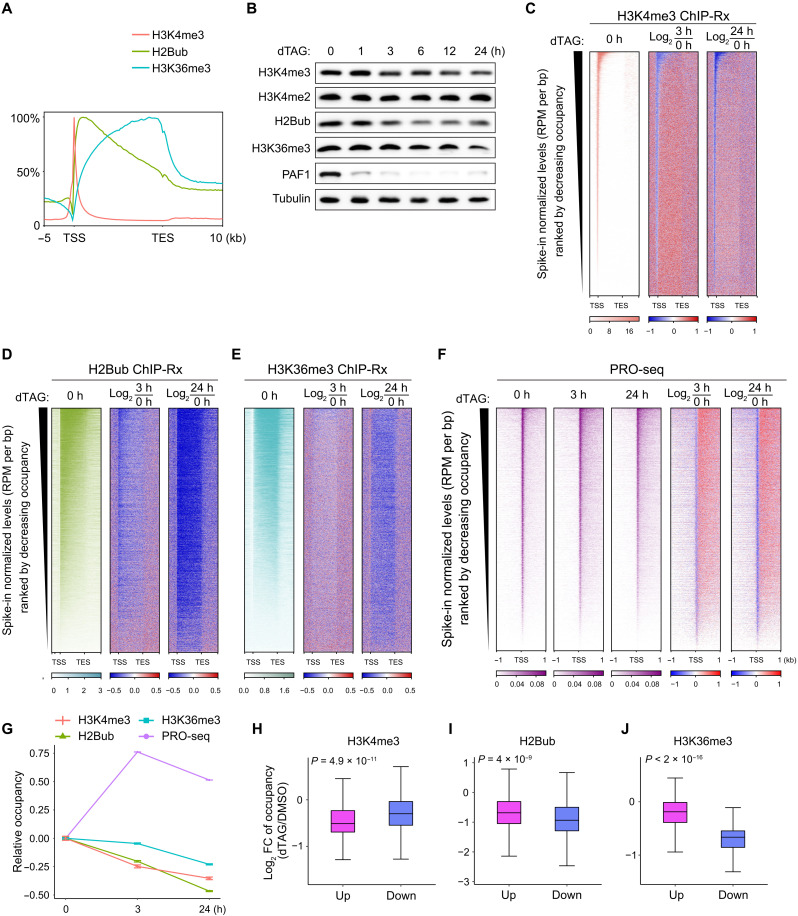
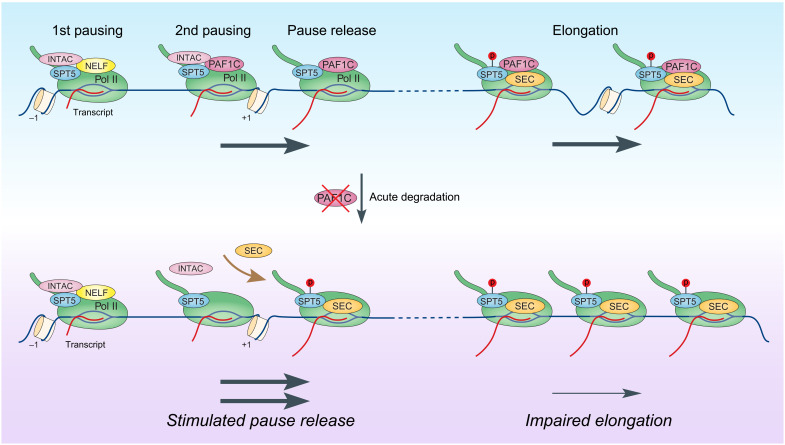
Pleiotropic transcription regulator RNA polymerase II (Pol II)-associated factor 1 (PAF1) governs multiple transcriptional steps and the deposition of several epigenetic marks. However, it remains unclear how ultimate transcriptional outcome is determined by PAF1 and whether it relates to PAF1-controlled epigenetic marks. We use rapid degradation systems and reveal direct PAF1 functions in governing pausing partially by recruiting Integrator-PP2A (INTAC), in addition to ensuring elongation. Following acute PAF1 degradation, most destabilized polymerase undergoes effective release, which presumably relies on skewed balance between INTAC and P-TEFb, resulting in hyperphosphorylated substrates including SPT5. Impaired Pol II progression during elongation, along with altered pause release frequency, determines the final transcriptional outputs. Moreover, PAF1 degradation causes a cumulative decline in histone modifications. These epigenetic alterations in chromatin likely further influence the production of transcripts from PAF1 target genes.
Figures
Similar articles
-
SPT5 stabilizes RNA polymerase II, orchestrates transcription cycles, and maintains the enhancer landscape.Mol Cell. 2021 Nov 4;81(21):4425-4439.e6. doi: 10.1016/j.molcel.2021.08.029. Epub 2021 Sep 16. Mol Cell. 2021. PMID: 34534457
-
Paf1C regulates RNA polymerase II progression by modulating elongation rate.Proc Natl Acad Sci U S A. 2019 Jul 16;116(29):14583-14592. doi: 10.1073/pnas.1904324116. Epub 2019 Jun 27. Proc Natl Acad Sci U S A. 2019. PMID: 31249142 Free PMC article.
-
PAF1 regulation of promoter-proximal pause release via enhancer activation.Science. 2017 Sep 22;357(6357):1294-1298. doi: 10.1126/science.aan3269. Epub 2017 Aug 31. Science. 2017. PMID: 28860207 Free PMC article.
-
The pleiotropic roles of SPT5 in transcription.Transcription. 2022 Feb-Jun;13(1-3):53-69. doi: 10.1080/21541264.2022.2103366. Epub 2022 Jul 25. Transcription. 2022. PMID: 35876486 Free PMC article. Review.
-
PD2/PAF1 at the Crossroads of the Cancer Network.Cancer Res. 2018 Jan 15;78(2):313-319. doi: 10.1158/0008-5472.CAN-17-2175. Epub 2018 Jan 8. Cancer Res. 2018. PMID: 29311159 Free PMC article. Review.
Cited by
-
YTHDC1 m6A-dependent and m6A-independent functions converge to preserve the DNA damage response.EMBO J. 2024 Aug;43(16):3494-3522. doi: 10.1038/s44318-024-00153-x. Epub 2024 Jul 1. EMBO J. 2024. PMID: 38951610 Free PMC article.
-
NusG-Spt5 Transcription Factors: Universal, Dynamic Modulators of Gene Expression.J Mol Biol. 2025 Jan 1;437(1):168814. doi: 10.1016/j.jmb.2024.168814. Epub 2024 Oct 5. J Mol Biol. 2025. PMID: 39374889 Free PMC article. Review.
-
Nuclear RNA catabolism controls endogenous retroviruses, gene expression asymmetry, and dedifferentiation.Mol Cell. 2023 Dec 7;83(23):4255-4271.e9. doi: 10.1016/j.molcel.2023.10.036. Epub 2023 Nov 22. Mol Cell. 2023. PMID: 37995687 Free PMC article.
-
Spt6 directly interacts with Cdc73 and is required for Paf1 complex occupancy at active genes in Saccharomyces cerevisiae.Nucleic Acids Res. 2023 Jun 9;51(10):4814-4830. doi: 10.1093/nar/gkad180. Nucleic Acids Res. 2023. PMID: 36928138 Free PMC article.
-
CDK12 and Integrator-PP2A complex modulates LEO1 phosphorylation for processive transcription elongation.Sci Adv. 2023 May 19;9(20):eadf8698. doi: 10.1126/sciadv.adf8698. Epub 2023 May 19. Sci Adv. 2023. PMID: 37205756 Free PMC article.
References
-
- Chen F. X., Smith E. R., Shilatifard A., Born to run: Control of transcription elongation by RNA polymerase II. Nat. Rev. Mol. Cell Biol. 19, 464–478 (2018). - PubMed
-
- Cramer P., Organization and regulation of gene transcription. Nature 573, 45–54 (2019). - PubMed
-
- Noe Gonzalez M., Blears D., Svejstrup J. Q., Causes and consequences of RNA polymerase II stalling during transcript elongation. Nat. Rev. Mol. Cell Biol. 22, 3–21 (2021). - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
