

Specification of fetal liver endothelial progenitors to functional zonated adult sinusoids requires c-Maf induction
- PMID: 35364013
- PMCID: PMC9290393
- DOI: 10.1016/j.stem.2022.03.002
Specification of fetal liver endothelial progenitors to functional zonated adult sinusoids requires c-Maf induction
Abstract
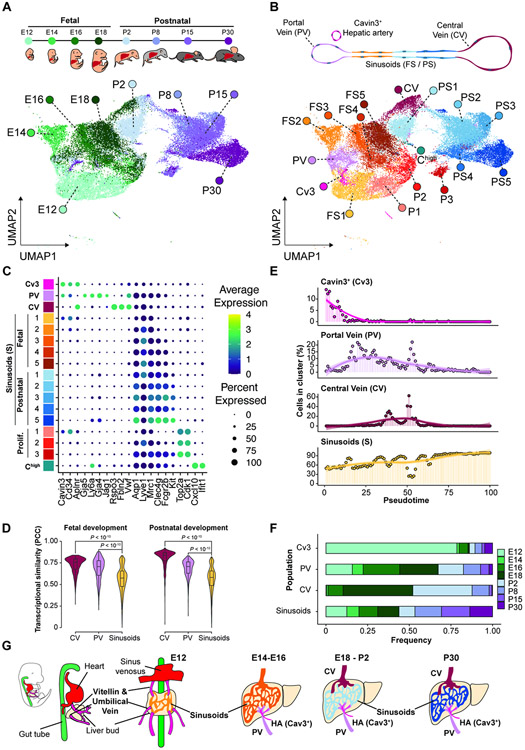
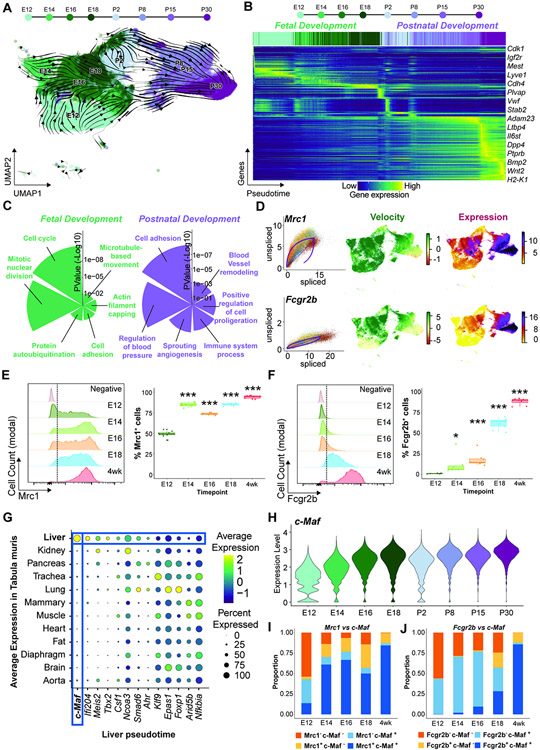
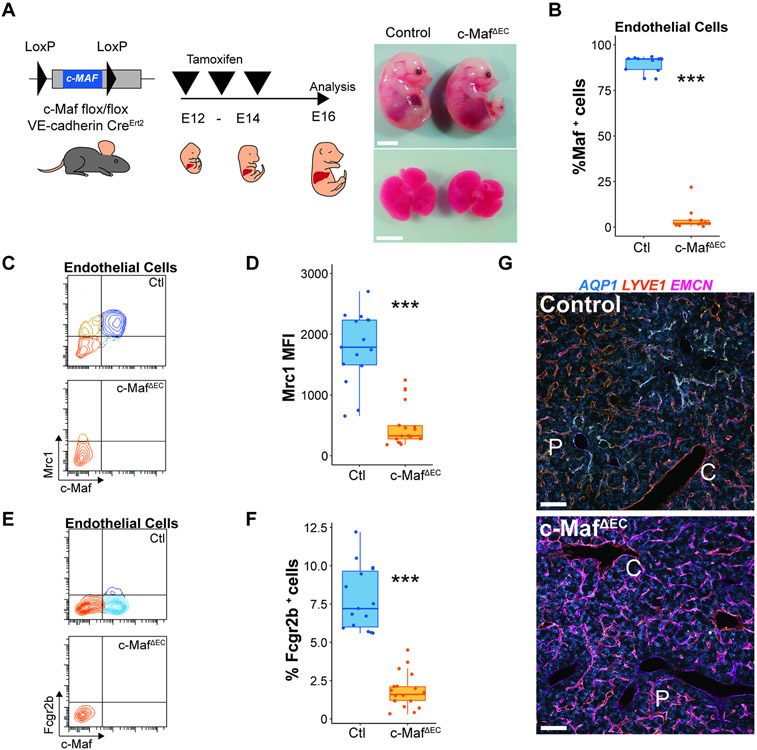
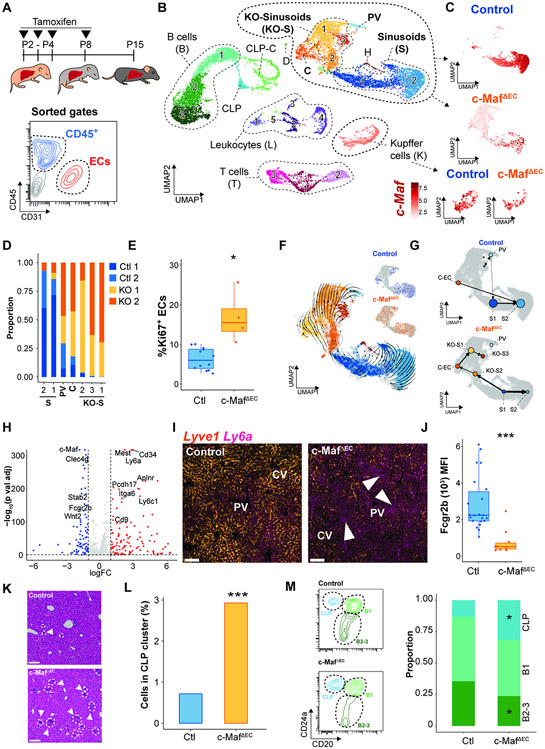
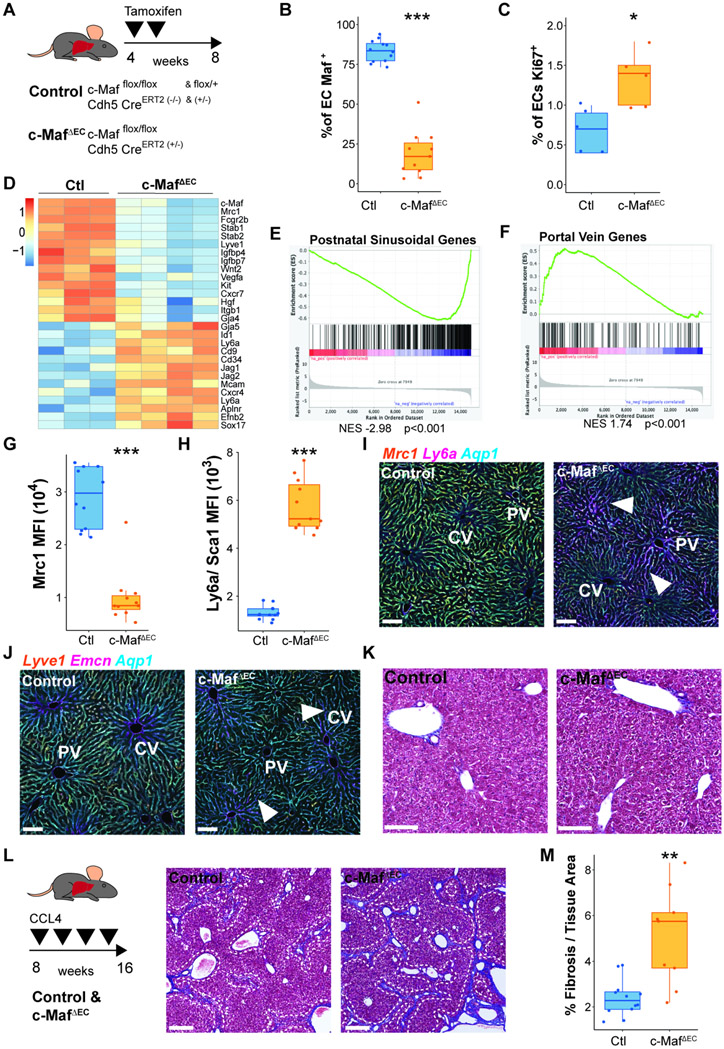
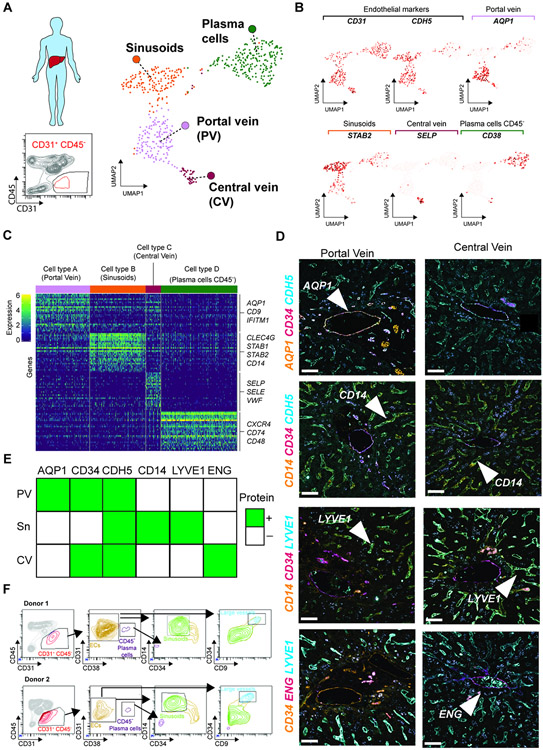
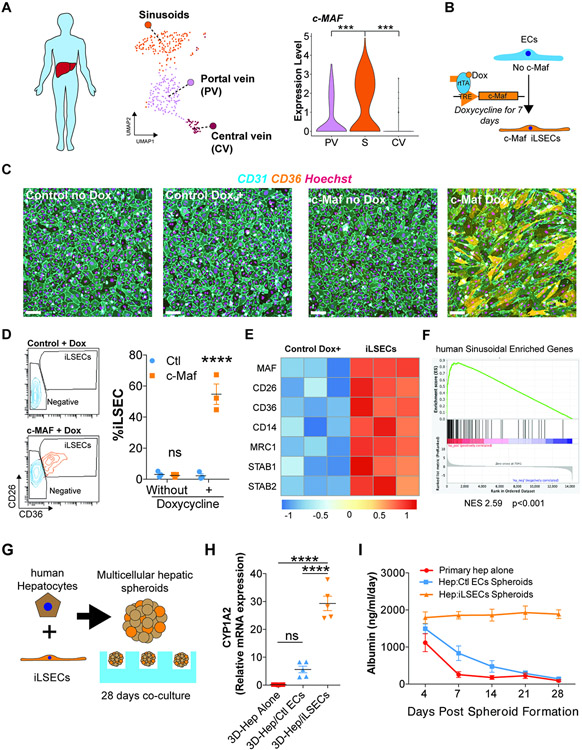
The liver vascular network is patterned by sinusoidal and hepatocyte co-zonation. How intra-liver vessels acquire their hierarchical specialized functions is unknown. We study heterogeneity of hepatic vascular cells during mouse development through functional and single-cell RNA-sequencing. The acquisition of sinusoidal endothelial cell identity is initiated during early development and completed postnatally, originating from a pool of undifferentiated vascular progenitors at E12. The peri-natal induction of the transcription factor c-Maf is a critical switch for the sinusoidal identity determination. Endothelium-restricted deletion of c-Maf disrupts liver sinusoidal development, aberrantly expands postnatal liver hematopoiesis, promotes excessive postnatal sinusoidal proliferation, and aggravates liver pro-fibrotic sensitivity to chemical insult. Enforced c-Maf overexpression in generic human endothelial cells switches on a liver sinusoidal transcriptional program that maintains hepatocyte function. c-Maf represents an inducible intra-organotypic and niche-responsive molecular determinant of hepatic sinusoidal cell identity and lays the foundation for the strategies for vasculature-driven liver repair.
Keywords: c-Maf; development; endothelial cell reprogramming; endothelial cell specification; fibrosis; hepatic angiocrine factors; liver sinusoidal endothelial cells; postnatal maturation; single-cell RNAseq; single-cell molecular profiling; vascular heterogeneity.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S. Rafii is the co-founder and non-paid consultant to Angiocrine Bioscience, San Diego, CA. R.E.S. is on the scientific advisory board of Miromatrix Inc. and is a speaker and consultant for Alnylam Inc.
Figures
Comment in
-
Endothelial cell diversity in the liver.Nat Rev Mol Cell Biol. 2022 May;23(5):305. doi: 10.1038/s41580-022-00481-y. Nat Rev Mol Cell Biol. 2022. PMID: 35383337 No abstract available.
-
c-Maf: The magic wand that turns on LSEC fate.Cell Stem Cell. 2022 Apr 7;29(4):491-493. doi: 10.1016/j.stem.2022.03.011. Cell Stem Cell. 2022. PMID: 35395181
References
-
- Augustin HG, and Koh GY (2017). Organotypic vasculature: From descriptive heterogeneity to functional pathophysiology. Science 357. - PubMed
-
- Bankston PW, and Pino RM (1980). The development of the sinusoids of fetal rat liver: morphology of endothelial cells, Kupffer cells, and the transmural migration of blood cells into the sinusoids. Am. J. Anat 159, 1–15. - PubMed
-
- Barberá-Guillem E, Arrue JM, Ballesteros J, and Vidal-Vanaclocha F (1986). Structural changes in endothelial cells of developing rat liver in the transition from fetal to postnatal life. J. Ultrastruct. Mol. Struct. Res 97, 197–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
