

Mechanisms underlying ubiquitin-driven selective mitochondrial and bacterial autophagy
- PMID: 35364016
- PMCID: PMC9254164
- DOI: 10.1016/j.molcel.2022.03.012
Mechanisms underlying ubiquitin-driven selective mitochondrial and bacterial autophagy
Abstract
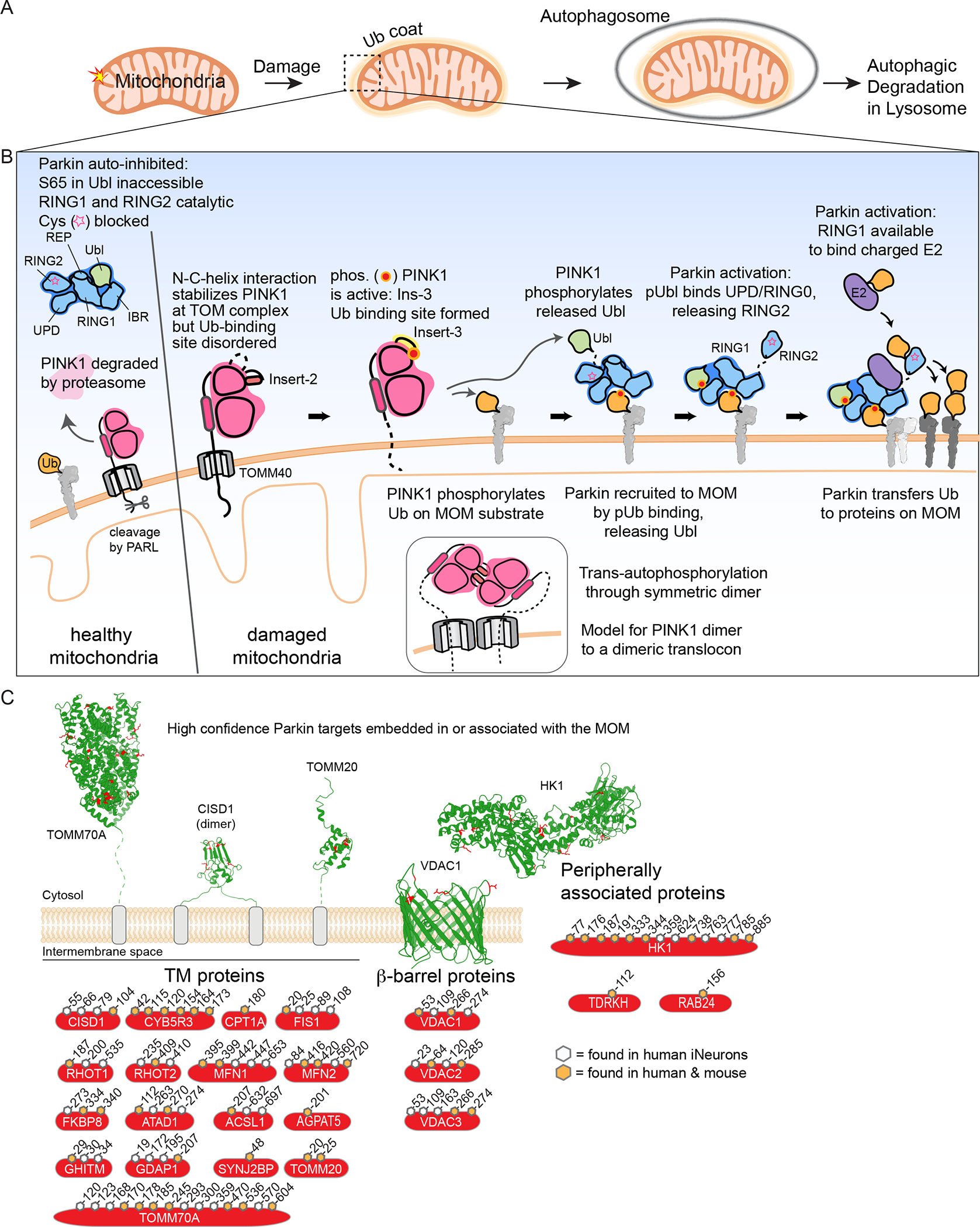
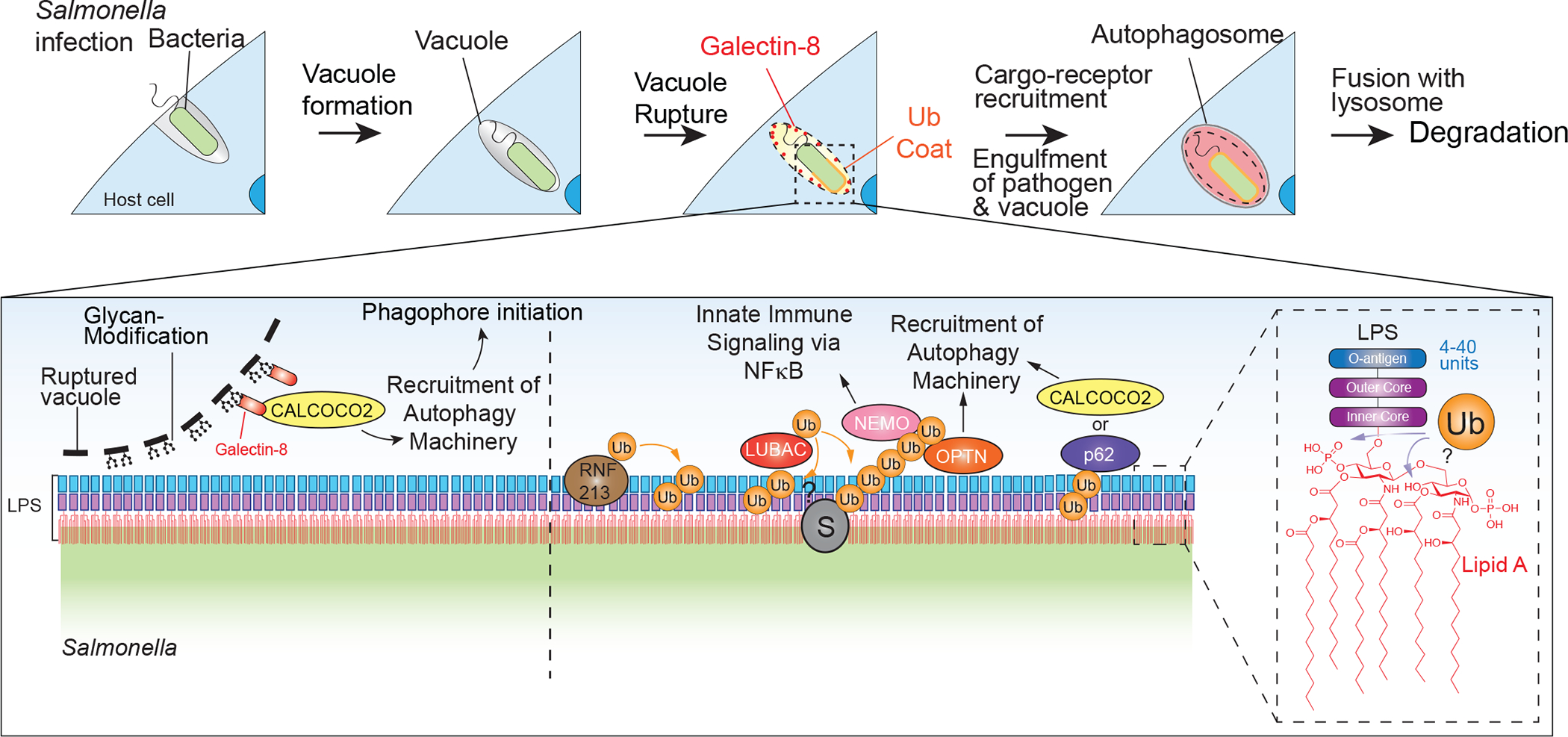
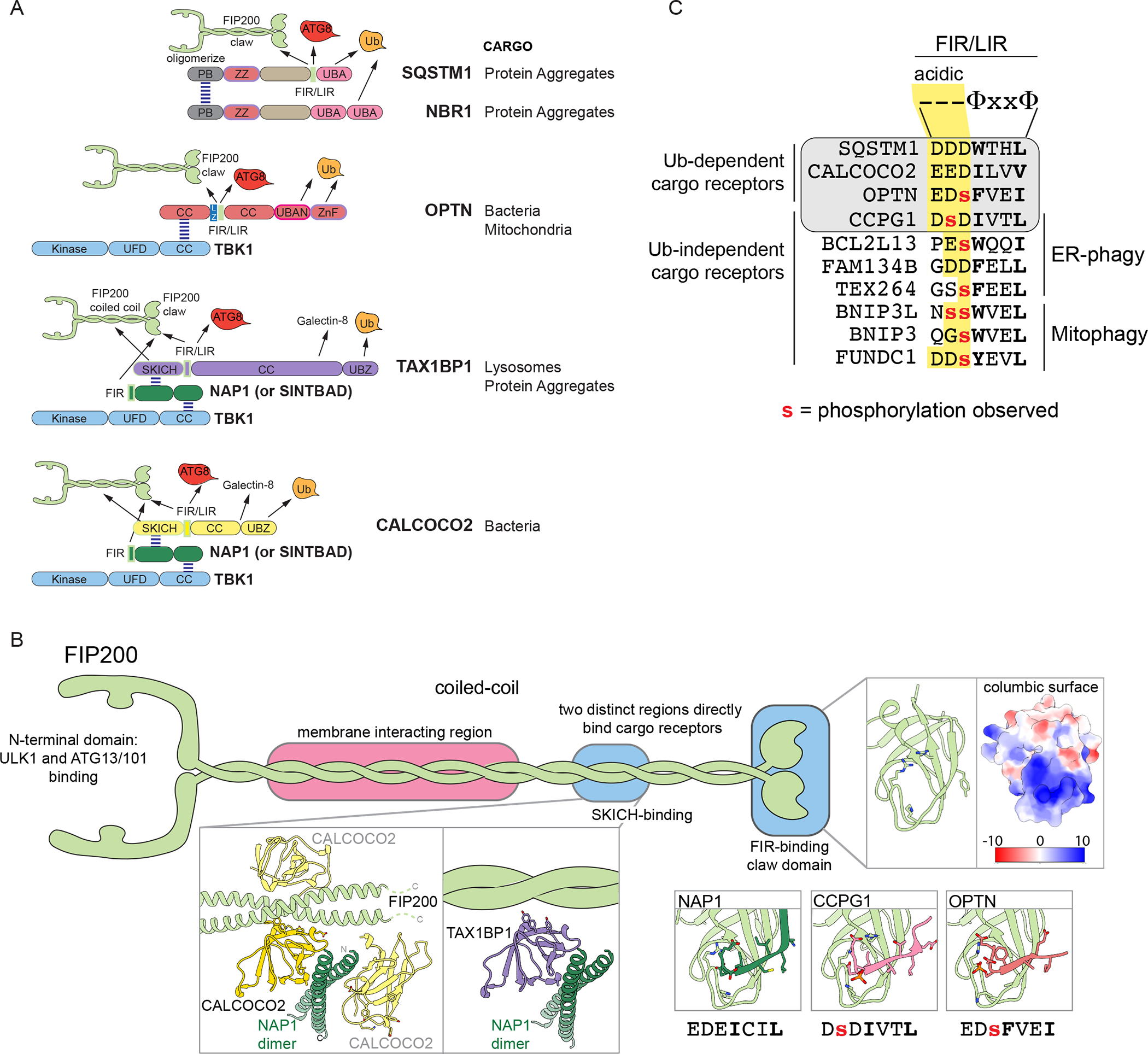
Selective autophagy specifically eliminates damaged or superfluous organelles, maintaining cellular health. In this process, a double membrane structure termed an autophagosome captures target organelles or proteins and delivers this cargo to the lysosome for degradation. The attachment of the small protein ubiquitin to cargo has emerged as a common mechanism for initiating organelle or protein capture by the autophagy machinery. In this process, a suite of ubiquitin-binding cargo receptors function to initiate autophagosome assembly in situ on the target cargo, thereby providing selectivity in cargo capture. Here, we review recent efforts to understand the biochemical mechanisms and principles by which cargo are marked with ubiquitin and how ubiquitin-binding cargo receptors use conserved structural modules to recruit the autophagosome initiation machinery, with a particular focus on mitochondria and intracellular bacteria as cargo. These emerging mechanisms provide answers to long-standing questions in the field concerning how selectivity in cargo degradation is achieved.
Keywords: cargo receptor; mitophagy; selective autophagy; ubiquitin; xenophagy.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.W.H. is a consultant and founder of Caraway Therapeutics and is a founding board member of Interline Therapeutics.
Figures

References
-
- Bingol B, Tea JS, Phu L, Reichelt M, Bakalarski CE, Song Q, Foreman O, Kirkpatrick DS, and Sheng M (2014). The mitochondrial deubiquitinase USP30 opposes parkin-mediated mitophagy. Nature 510, 370–375. - PubMed
-
- Boyle KB, and Randow F (2013). The role of ‘eat-me’ signals and autophagy cargo receptors in innate immunity. Curr Opin Microbiol 16, 339–348. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
