

Donor-Acceptor Pyridinium Salts for Photo-Induced Electron-Transfer-Driven Modification of Tryptophan in Peptides, Proteins, and Proteomes Using Visible Light
- PMID: 35364811
- PMCID: PMC10124759
- DOI: 10.1021/jacs.1c10536
Donor-Acceptor Pyridinium Salts for Photo-Induced Electron-Transfer-Driven Modification of Tryptophan in Peptides, Proteins, and Proteomes Using Visible Light
Abstract
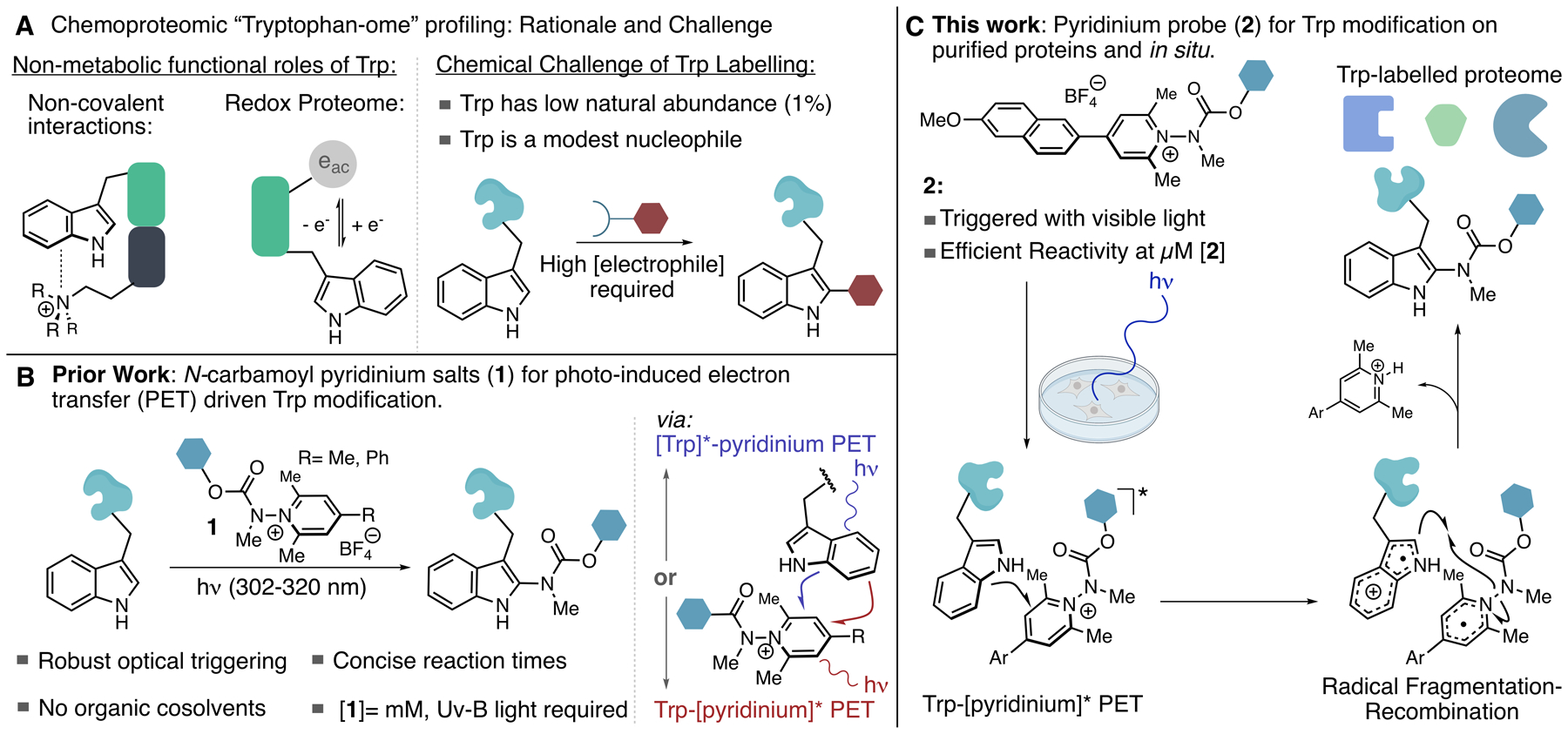
Tryptophan (Trp) plays a variety of critical functional roles in protein biochemistry; however, owing to its low natural frequency and poor nucleophilicity, the design of effective methods for both single protein bioconjugation at Trp as well as for in situ chemoproteomic profiling remains a challenge. Here, we report a method for covalent Trp modification that is suitable for both scenarios by invoking photo-induced electron transfer (PET) as a means of driving efficient reactivity. We have engineered biaryl N-carbamoyl pyridinium salts that possess a donor-acceptor relationship that enables optical triggering with visible light whilst simultaneously attenuating the probe's photo-oxidation potential in order to prevent photodegradation. This probe was assayed against a small bank of eight peptides and proteins, where it was found that micromolar concentrations of the probe and short irradiation times (10-60 min) with violet light enabled efficient reactivity toward surface exposed Trp residues. The carbamate transferring group can be used to transfer useful functional groups to proteins including affinity tags and click handles. DFT calculations and other mechanistic analyses reveal correlations between excited state lifetimes, relative fluorescence quantum yields, and chemical reactivity. Biotinylated and azide-functionalized pyridinium salts were used for Trp profiling in HEK293T lysates and in situ in HEK293T cells using 440 nm LED irradiation. Peptide-level enrichment from live cell labeling experiments identified 290 Trp modifications, with 82% selectivity for Trp modification over other π-amino acids, demonstrating the ability of this method to identify and quantify reactive Trp residues from live cells.
Conflict of interest statement
The authors declare the following competing financial interest(s): FJG, DTB, GEM, TRB, CHH MDJ, MS are employees of Novartis Institutes for BioMedical Research.
Figures






Similar articles
-
Selective Modification of Tryptophan Residues in Peptides and Proteins Using a Biomimetic Electron Transfer Process.J Am Chem Soc. 2020 May 20;142(20):9112-9118. doi: 10.1021/jacs.0c03039. Epub 2020 May 5. J Am Chem Soc. 2020. PMID: 32348670 Free PMC article.
-
Oxidative cyclization reagents reveal tryptophan cation-π interactions.Nature. 2024 Mar;627(8004):680-687. doi: 10.1038/s41586-024-07140-6. Epub 2024 Mar 6. Nature. 2024. PMID: 38448587 Free PMC article.
-
Photochemical Chemoselective Alkylation of Tryptophan-Containing Peptides.Org Lett. 2021 Jan 15;23(2):285-289. doi: 10.1021/acs.orglett.0c03735. Epub 2021 Jan 5. Org Lett. 2021. PMID: 33400540
-
N-Functionalized Pyridinium Salts: A New Chapter for Site-Selective Pyridine C-H Functionalization via Radical-Based Processes under Visible Light Irradiation.Acc Chem Res. 2022 Oct 18;55(20):3043-3056. doi: 10.1021/acs.accounts.2c00530. Epub 2022 Sep 27. Acc Chem Res. 2022. PMID: 36166489 Review.
-
Novel chemical degradation pathways of proteins mediated by tryptophan oxidation: tryptophan side chain fragmentation.J Pharm Pharmacol. 2018 May;70(5):655-665. doi: 10.1111/jphp.12688. Epub 2017 Jan 30. J Pharm Pharmacol. 2018. PMID: 28134972 Review.
Cited by
-
Clickable tryptophan modification for late-stage diversification of native peptides.Sci Adv. 2024 Jul 12;10(28):eadp9958. doi: 10.1126/sciadv.adp9958. Epub 2024 Jul 10. Sci Adv. 2024. PMID: 38985871 Free PMC article.
-
Redox-neutral, metal-free tryptophan labeling of polypeptides in hexafluoroisopropanol (HFIP).RSC Chem Biol. 2024 Aug 26;5(10):963-9. doi: 10.1039/d4cb00142g. Online ahead of print. RSC Chem Biol. 2024. PMID: 39234575 Free PMC article.
-
Chemoselective Late-Stage Functionalization of Peptides via Photocatalytic C2-Alkylation of Tryptophan.Org Lett. 2023 Jul 28;25(29):5459-5464. doi: 10.1021/acs.orglett.3c01795. Epub 2023 Jul 18. Org Lett. 2023. PMID: 37462428 Free PMC article.
-
Using dimethylsulfonium to identify readers of methylation.Nat Chem. 2024 Aug;16(8):1221-1222. doi: 10.1038/s41557-024-01582-1. Nat Chem. 2024. PMID: 39079946 No abstract available.
-
Photoredox catalysis enabling decarboxylative radical cyclization of γ,γ-dimethylallyltryptophan (DMAT) derivatives: formal synthesis of 6,7-secoagroclavine.Beilstein J Org Chem. 2023 Jun 26;19:918-927. doi: 10.3762/bjoc.19.70. eCollection 2023. Beilstein J Org Chem. 2023. PMID: 37404801 Free PMC article.
References
-
- Niphakis MJ; Cravatt BF Enzyme Inhibitor Discovery by Activity-Based Protein Profiling. Annu. Rev. Biochem 2014, 83, 341–77. - PubMed
- Cravatt BF; Wright AT; Kozarich JW Activity-Based Protein Profiling: From Enzyme Chemistry to Proteomic Chemistry. Annu Rev Biochem. 2008, 77, 383–414. - PubMed
- Heal WP; Dang. THT; Tate EW Activity-based probes: discovering new biology and new drug targets. Chem Soc Rev. 2011, 40, 246–57. - PubMed
- Chan WC; Sharifzadeh S; Buhrlage SJ; Marto JA Chemoproteomic methods for covalent drug discovery. Chem. Soc. Rev 2021, 50, 8361–8381. - PMC - PubMed
- Spradlin JN; Zhang E; Nomura DK Reimagining Druggability Using Chemoproteomic Platforms. Acc. Chem. Res 2021, 54, 1801–1813. - PubMed
-
-
For reviews summarizing cysteine profiling:
- Maurais AJ; Weerapana E Reactive-cysteine profiling for drug discovery. Curr. Op. Chem. Biol 2019, 50, 29–36. - PMC - PubMed
- Backus KM; Applications of Reactive Cysteine Profiling. Curr. Top. Microbiol. Immunol 2019, 420, 375–417. - PubMed
- Guan I; Williams K; Pan J; Liu X New Cysteine Covalent Modification Strategies Enable Advancement of Proteome-wide Selectivity of Kinase Modulators. Asian J. Org. Chem 2021, 10, 949–963.
-
For examples of lysine profiling
- Hacker SM; Backus KM; Lazear MR; Forli S; Correia BE; Cravatt BF Global profiling of lysine reactivity and ligandability in the human proteome. Nature Chemistry. 2017, 9, 1181–1190. - PMC - PubMed
- Shannon DA; Banerjee R; Webster ER; Bak DW; Wang C; Weerapana E Investigating the Proteome Reactivity and Selectivity of Aryl Halides. J. Am. Chem. Soc 2014, 136, 3330–3333. - PubMed
-
For examples targeting N-, O- and S-nucleophiles:
- Ward CC; Kleinman JI; Nomura DK NHS-Esters As Versatile Reactivity-Based Probes for Mapping Proteome-Wide Ligandable Hotspots. ACS Chem. Biol 2017, 12, 1478–1483. - PMC - PubMed
-
For examples of Tyrosine profiling:
- Brulet JW; Borne AL; Yuan K; Libby AH; Hsu K-L Liganding Functional Tyrosine Sites on Proteins Using Sulfur-Triazole Exchange Chemistry. J. Am. Chem. Soc 2020, 142, 18, 8270–8280. - PMC - PubMed
- Sun F; Suttapitugsakul S; Wu R An Azo Coupling-Based Chemoproteomic Approach to Systematically Profile the Tyrosine Reactivity in the Human Proteome. Anal. Chem 2021, 93, 10334–10342. - PMC - PubMed
-
For Carboxylic acid profiling:
- Bach K; Beerkens BLH; Zanon PRA; Hacker SM Light-Activatable, 2,5-Disubstituted Tetrazoles for the Proteome-wide Profiling of Aspartates and Glutamates in Living Bacteria. ACS. Cent. Sci 2020, 6, 546–554. - PMC - PubMed
- Ma N; Hu J; Zhang Z-M; Liu W; Huang M; Fan. Y; Yin X; Wang J; Ding K; Ye W; Li Z 2H-Azirine-Based Reagents for Chemoselective Bioconjugation Carboxyl Residues Inside Live Cells. J. Am. Chem. Soc 2020, 142, 6051–6059. - PubMed
-
For Methionine profiling:
- Lin S; Yang X; Jia S; Weeks AM; Hornsby M; Lee PS; Nichiporuk RV; Iavarone AT; Wells JA; Toste FD; Chang CJ Redox-based reagents for chemoselective methionine bioconjugation. Science. 2017, 355, 597–602. - PMC - PubMed
-
-
- Lakhdar S; Westermaier M; Terrier R; Boubaker T; Ofial AR; Mayr H Nucleophilic Reactivites of Indoles. J. Org. Chem 2006, 71, 24, 9088–9095. - PubMed
-
- Scoffone E; Fontana A; Rocchi R Sulfenyl halides as Modifying Reagents for Polypeptides and Protein I. Modification of Tryptophan Residues. Biochemistry. 1968, 7, 971–979. - PubMed
- Kuyama H; Watanabe M; Toda C; Ando E; Tanaka K; Nishimura O An approach to quantitative proteome analysis by labeling tryptophan residues. Rapid Commun. Mass Spectrom 2003, 17, 1642–1650. - PubMed
- Antos JM; McFarland JM; Ivarone AT; Francis MB Chemoselective Tryptophan Labeling with Rhodium Carbenoids at Mild pH. J. Am. Chem. Soc 2009, 131, 6301–6308. - PMC - PubMed
- Ball ZT Designing Enzyme-like Catalysts: A Rhodium(II) Metallopeptide Case Study. Acc. Chem. Res 2013. 46, 560–570. - PubMed
- Seki Y; Ishiyama T; Sasaki D; Abe J; Sohma Y; Oisaka K; Kanai M Transition Metal-Free Tryptophan-Selective Bioconjugation of Proteins. J. Am. Chem. Soc 2016, 138, 10798–10801. - PubMed
- Wang W; Lorion MM; Shah J; Kapdi AR; Ackermann L Late-Stage Peptide Diversification by Position-Selective C–H activation. Angew. Chem. Int. Ed 2018, 57, 2–20. - PubMed
- Petersen J; Christensen KE; Nielsen MT; Mortensen KT; Komnatnyy VV; Nielsen TE; Qvortrup K Oxidative Modification of Tryptophan-Containing Peptides. ACS. Comb. Sci 2018, 20, 344–349. - PubMed
- Foettinger A; Leitner A; Lindner W Selective Enrichment of Trytophan-Containing Peptides from Protein Digests Employing a Reversible Derivatization with Malondialdehyde and Solid-Phase Capture on Hydrazide Beads. J. Proteome Res 2007, 6, 3827–3834. - PubMed
- Yu Y; Zhang LK; Buevich AV; Li G; Tang H; Vachal P; Colletti SL; Shi ZC Chemoselective peptide Modification via Photocataliyic Tryptophan β-position Conjugation. J. Am. Chem. Soc 2018, 140, 6797–6800. - PubMed
-
- Tsai C-J; Lin SL; Wolfson HJ; Nussinov R Studies of protein-protein interfaces: A statistical analysis of the hydrophobic effect. Protein Sci. 1997, 6, 53–64. - PMC - PubMed
- Ma B; Elkayam T; Wolfson H; Nussinov R Protein-protein interactions: Structurally conserved residues distinguish between binding sites and exposed protein surfaces. Proc. Natl. Acad. Sci 2003, 100, 5772–5777. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
