

Transsulfuration, minor player or crucial for cysteine homeostasis in cancer
- PMID: 35365367
- PMCID: PMC9378356
- DOI: 10.1016/j.tcb.2022.02.009
Transsulfuration, minor player or crucial for cysteine homeostasis in cancer
Abstract
Cysteine, a thiol-containing amino acid, is crucial for the synthesis of sulfur-containing biomolecules that control multiple essential cellular activities. Altered cysteine metabolism has been linked to numerous driver oncoproteins and tumor suppressors, as well as to malignant traits in cancer. Cysteine can be acquired from extracellular sources or synthesized de novo via the transsulfuration (TSS) pathway. Limited availability of cystine in tumor interstitial fluids raises the possible dependency on de novo cysteine synthesis via TSS. However, the contribution of TSS to cancer metabolism remains highly contentious. Based on recent findings, we provide new perspectives on this crucial but understudied metabolic pathway in cancer.
Keywords: cancer; cysteine metabolism; ferroptosis; glutathione; redox homeostasis; transsulfuration.
Copyright © 2022 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures
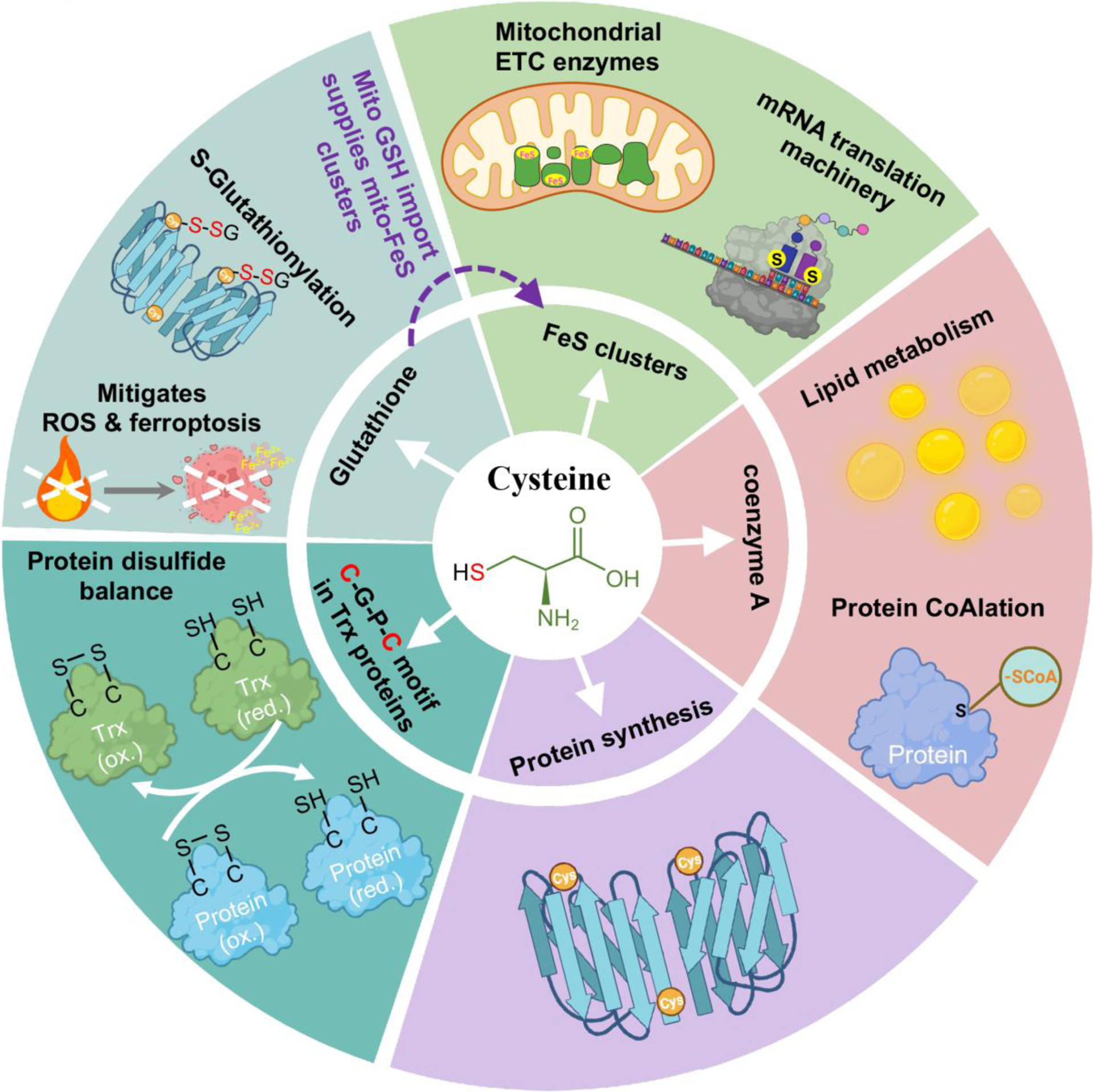
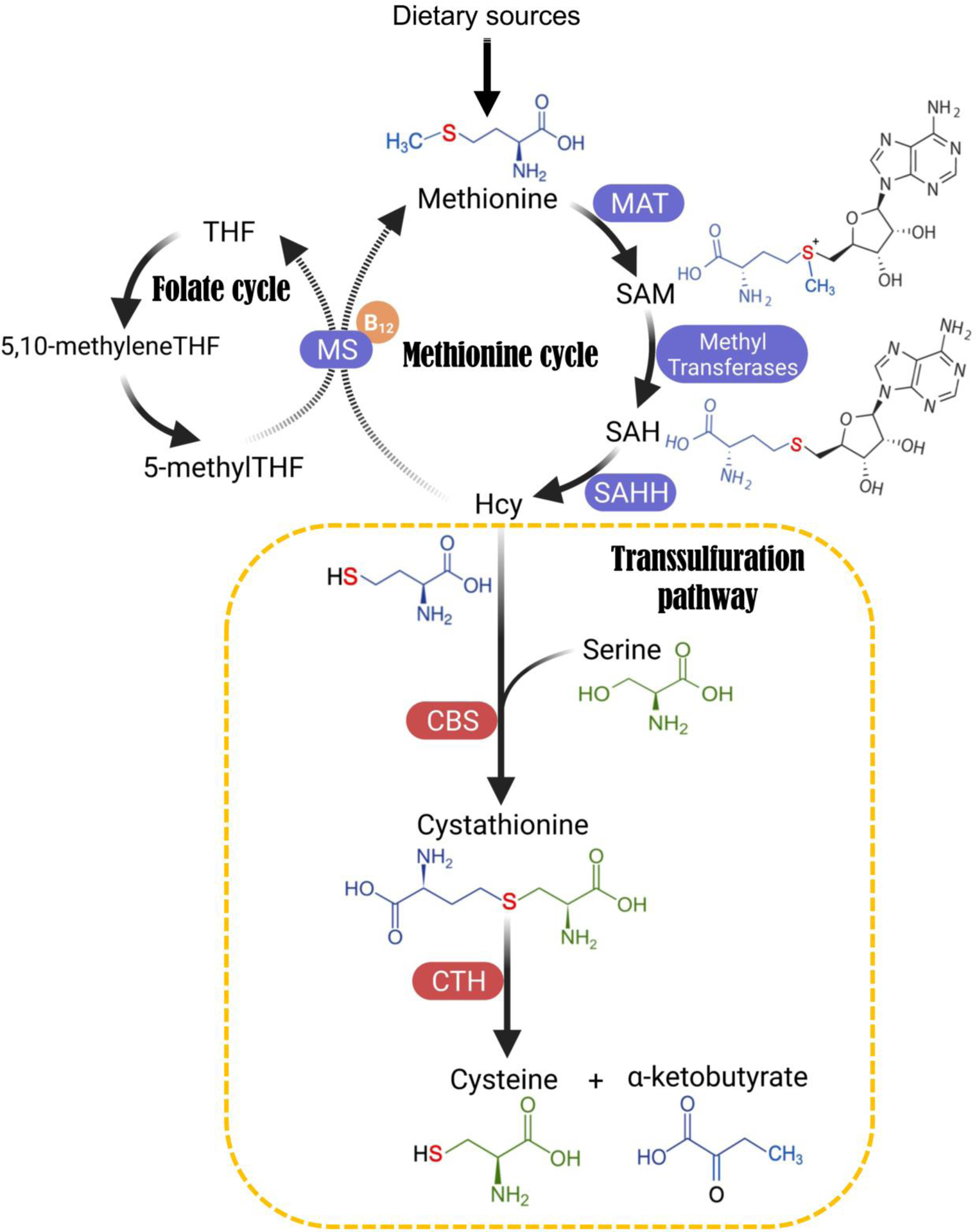
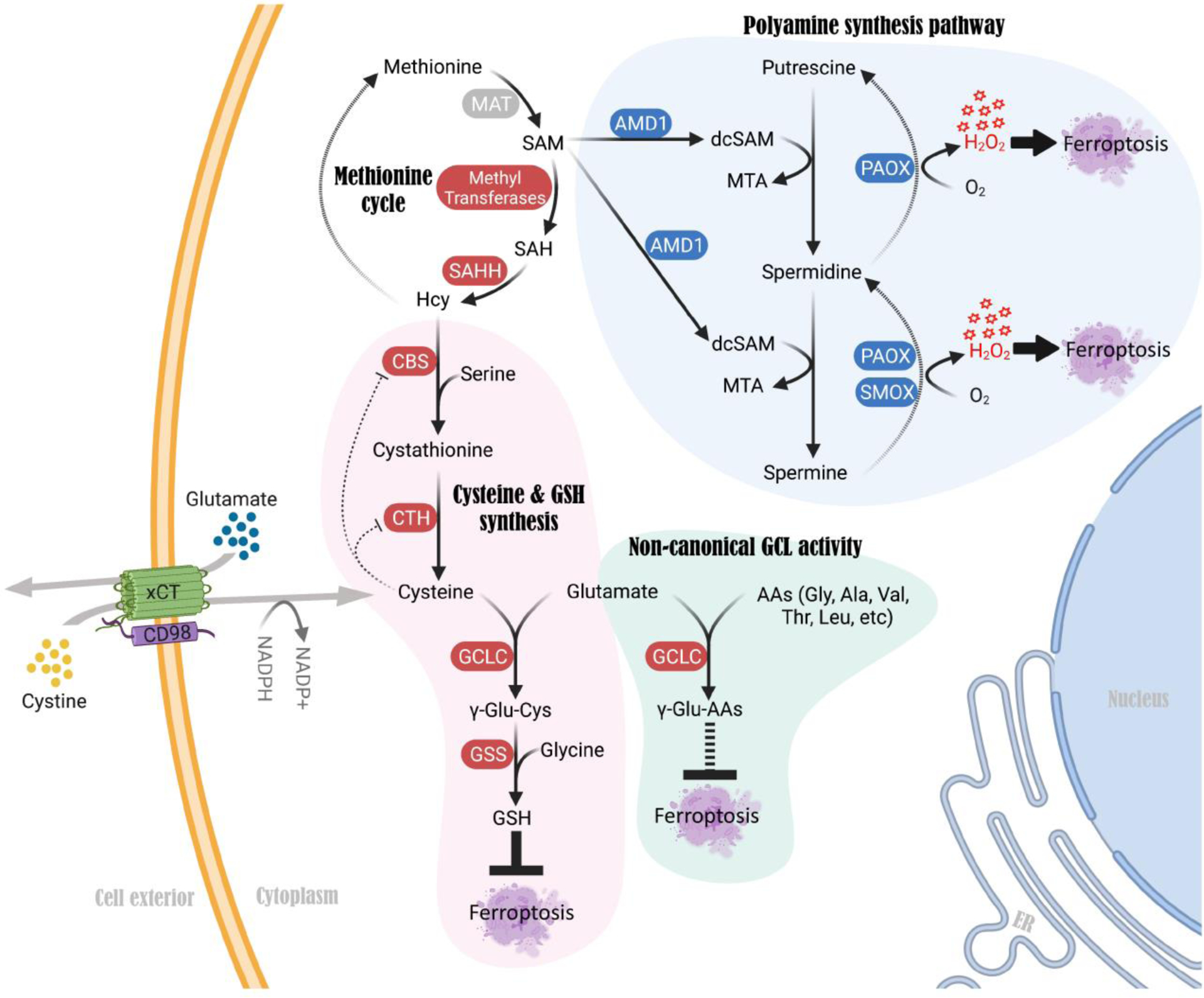
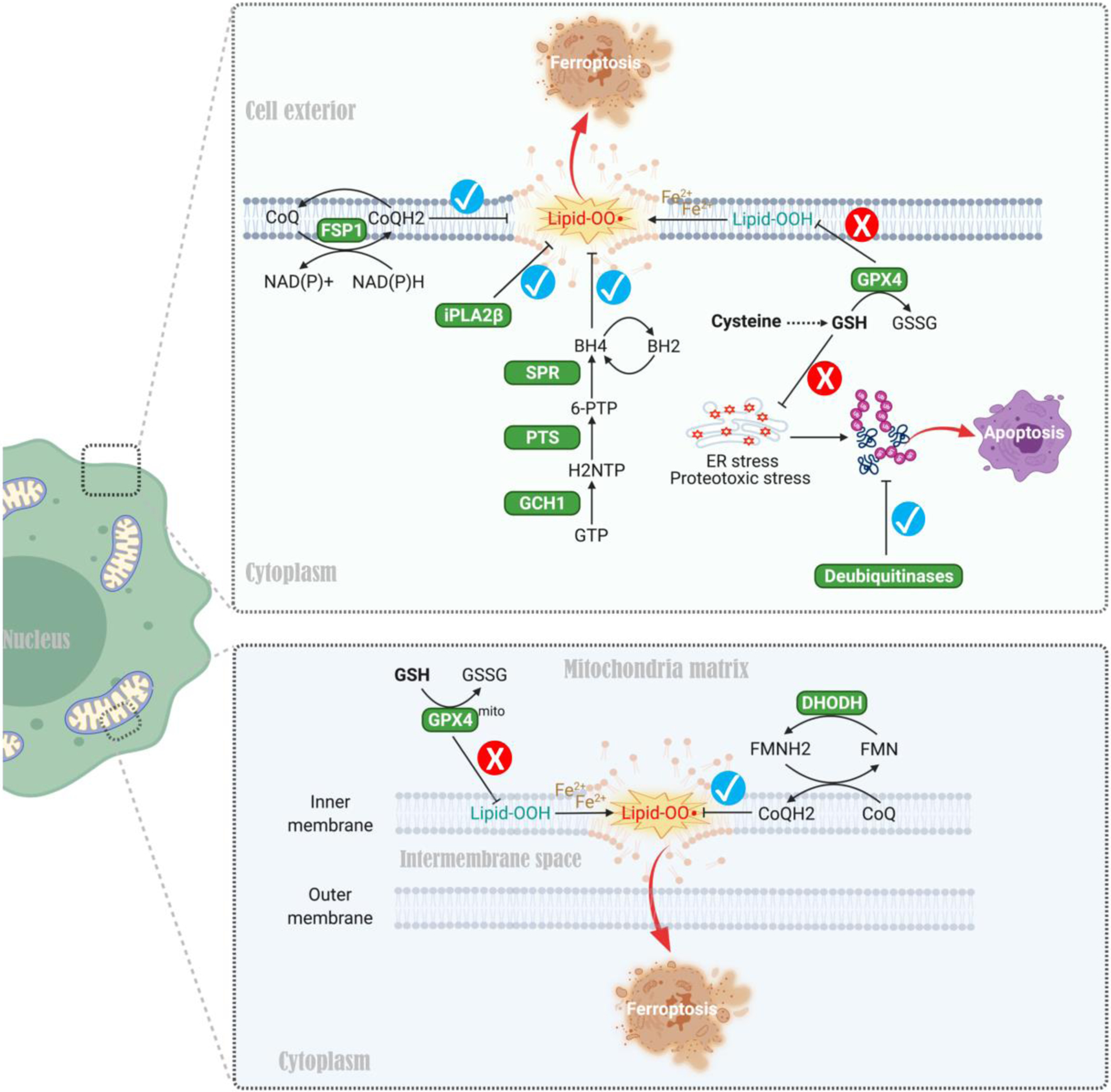
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
