

AF2Complex predicts direct physical interactions in multimeric proteins with deep learning
- PMID: 35365655
- PMCID: PMC8975832
- DOI: 10.1038/s41467-022-29394-2
AF2Complex predicts direct physical interactions in multimeric proteins with deep learning
Abstract
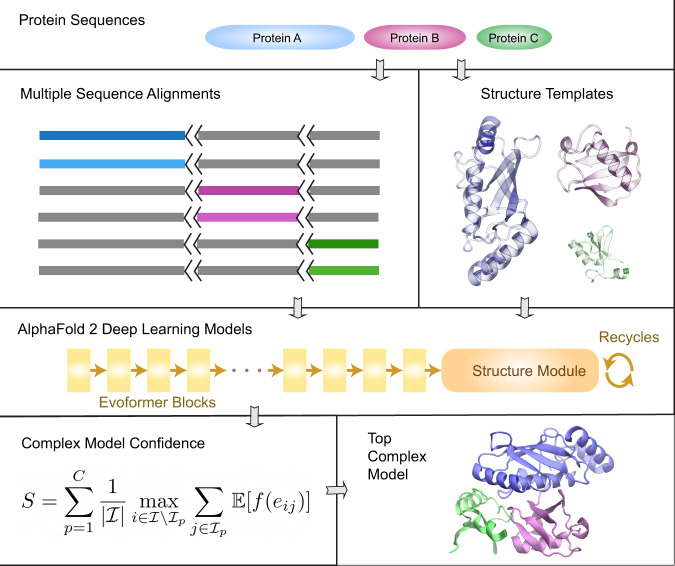
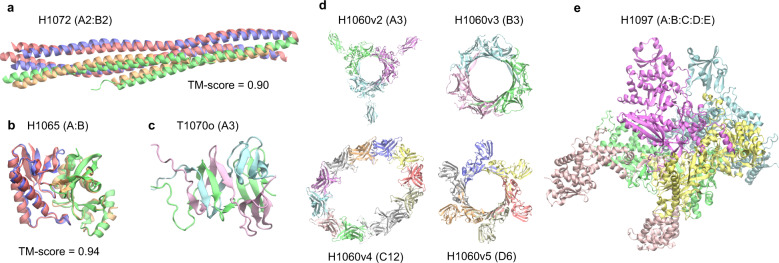
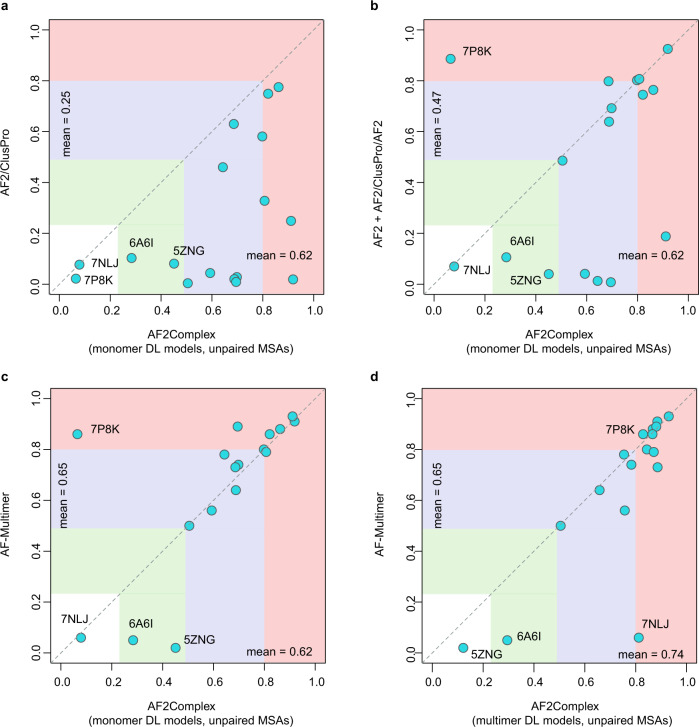
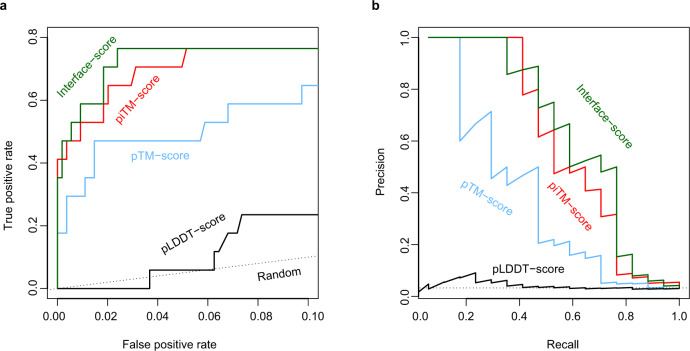
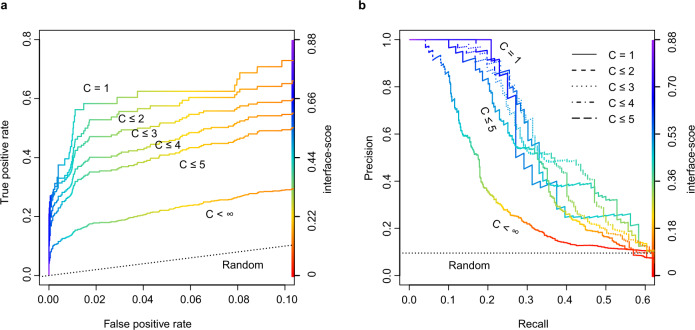
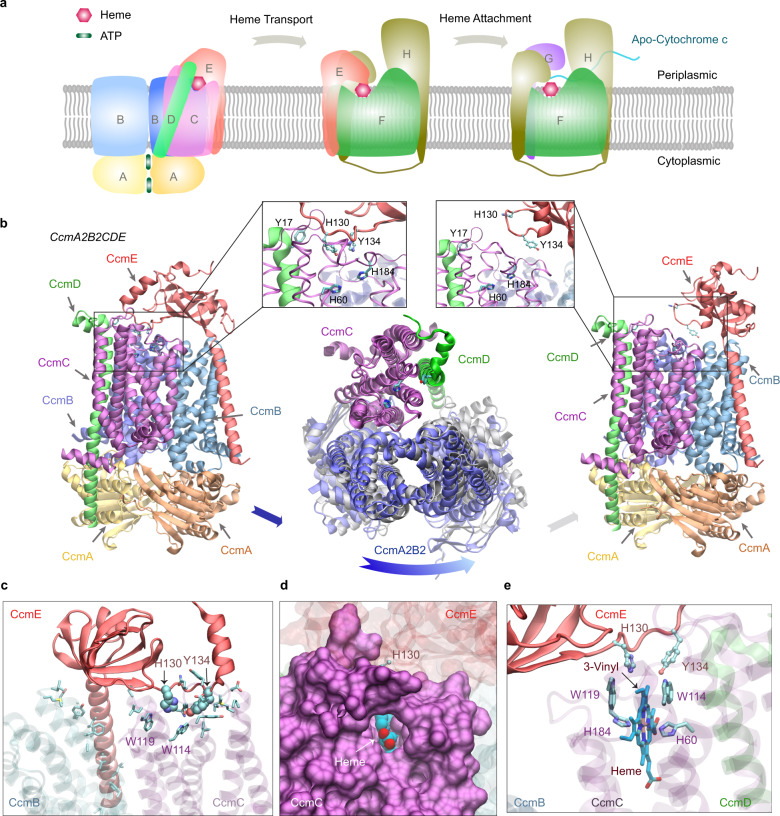
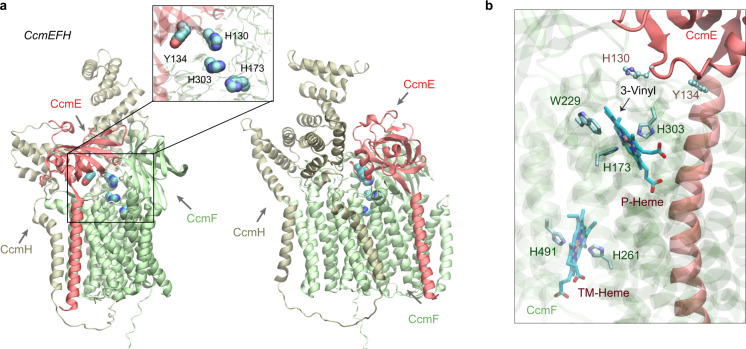
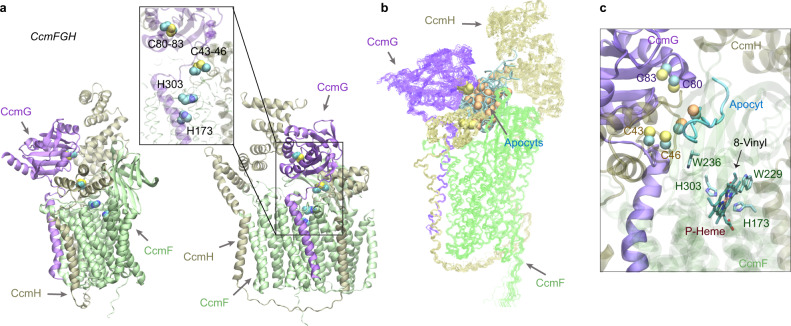
Accurate descriptions of protein-protein interactions are essential for understanding biological systems. Remarkably accurate atomic structures have been recently computed for individual proteins by AlphaFold2 (AF2). Here, we demonstrate that the same neural network models from AF2 developed for single protein sequences can be adapted to predict the structures of multimeric protein complexes without retraining. In contrast to common approaches, our method, AF2Complex, does not require paired multiple sequence alignments. It achieves higher accuracy than some complex protein-protein docking strategies and provides a significant improvement over AF-Multimer, a development of AlphaFold for multimeric proteins. Moreover, we introduce metrics for predicting direct protein-protein interactions between arbitrary protein pairs and validate AF2Complex on some challenging benchmark sets and the E. coli proteome. Lastly, using the cytochrome c biogenesis system I as an example, we present high-confidence models of three sought-after assemblies formed by eight members of this system.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Deep learning-driven insights into super protein complexes for outer membrane protein biogenesis in bacteria.Elife. 2022 Dec 28;11:e82885. doi: 10.7554/eLife.82885. Elife. 2022. PMID: 36576775 Free PMC article.
-
TopoQA: a topological deep learning-based approach for protein complex structure interface quality assessment.Brief Bioinform. 2025 Mar 4;26(2):bbaf083. doi: 10.1093/bib/bbaf083. Brief Bioinform. 2025. PMID: 40062613 Free PMC article.
-
ProALIGN: Directly Learning Alignments for Protein Structure Prediction via Exploiting Context-Specific Alignment Motifs.J Comput Biol. 2022 Feb;29(2):92-105. doi: 10.1089/cmb.2021.0430. Epub 2022 Jan 21. J Comput Biol. 2022. PMID: 35073170 Free PMC article.
-
A Perspective on the Prospective Use of AI in Protein Structure Prediction.J Chem Inf Model. 2024 Jan 8;64(1):26-41. doi: 10.1021/acs.jcim.3c01361. Epub 2023 Dec 20. J Chem Inf Model. 2024. PMID: 38124369 Review.
-
Multimeric protein interaction and complex prediction: Structure, dynamics and function.Comput Struct Biotechnol J. 2025 May 16;27:1975-1997. doi: 10.1016/j.csbj.2025.05.009. eCollection 2025. Comput Struct Biotechnol J. 2025. PMID: 40496891 Free PMC article. Review.
Cited by
-
The translocation assembly module (TAM) catalyzes the assembly of bacterial outer membrane proteins in vitro.Nat Commun. 2024 Aug 23;15(1):7246. doi: 10.1038/s41467-024-51628-8. Nat Commun. 2024. PMID: 39174534 Free PMC article.
-
Protein structure prediction in the era of AI: Challenges and limitations when applying to in silico force spectroscopy.Front Bioinform. 2022 Oct 7;2:983306. doi: 10.3389/fbinf.2022.983306. eCollection 2022. Front Bioinform. 2022. PMID: 36304287 Free PMC article.
-
Computational drug development for membrane protein targets.Nat Biotechnol. 2024 Feb;42(2):229-242. doi: 10.1038/s41587-023-01987-2. Epub 2024 Feb 15. Nat Biotechnol. 2024. PMID: 38361054 Review.
-
Prediction of protein structure and AI.J Hum Genet. 2024 Oct;69(10):477-480. doi: 10.1038/s10038-023-01215-4. Epub 2024 Jan 4. J Hum Genet. 2024. PMID: 38177398 Review.
-
Structurally-informed human interactome reveals proteome-wide perturbations by disease mutations.bioRxiv [Preprint]. 2024 Feb 1:2023.04.24.538110. doi: 10.1101/2023.04.24.538110. bioRxiv. 2024. Update in: Nat Biotechnol. 2024 Oct 24. doi: 10.1038/s41587-024-02428-4. PMID: 37162909 Free PMC article. Updated. Preprint.
References
-
- Marcotte EM, et al. Detecting Protein Function and Protein-Protein Interactions from Genome Sequences. Science. 1999;285:751–753. - PubMed
-
- Keskin Z, Gursoy A, Ma B, Nussinov R. Principles of protein-protein interactions: What are the preferred ways for proteins to interact? Chem. Rev. 2008;108:1225–1244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
