

Dietary fat promotes antibiotic-induced Clostridioides difficile mortality in mice
- PMID: 35365681
- PMCID: PMC8975876
- DOI: 10.1038/s41522-022-00276-1
Dietary fat promotes antibiotic-induced Clostridioides difficile mortality in mice
Abstract
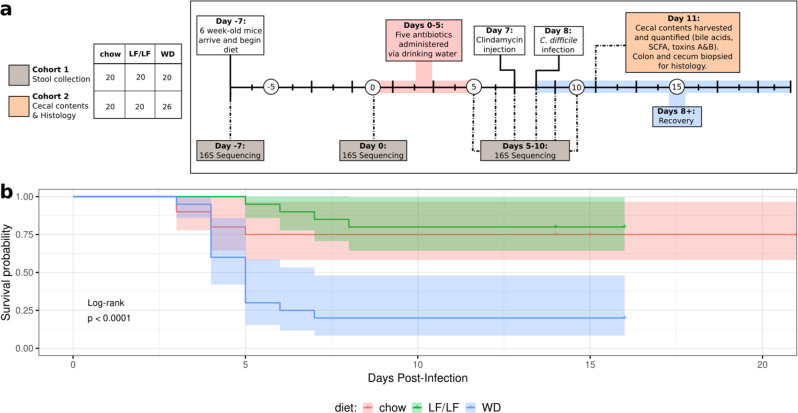
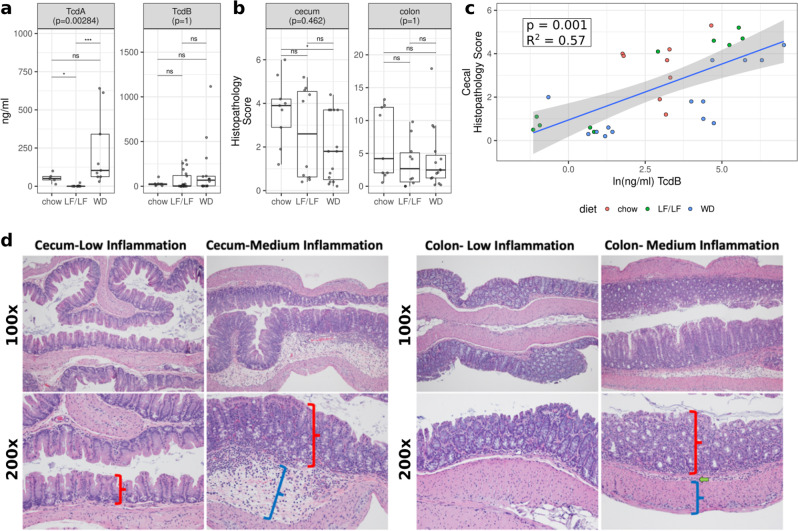
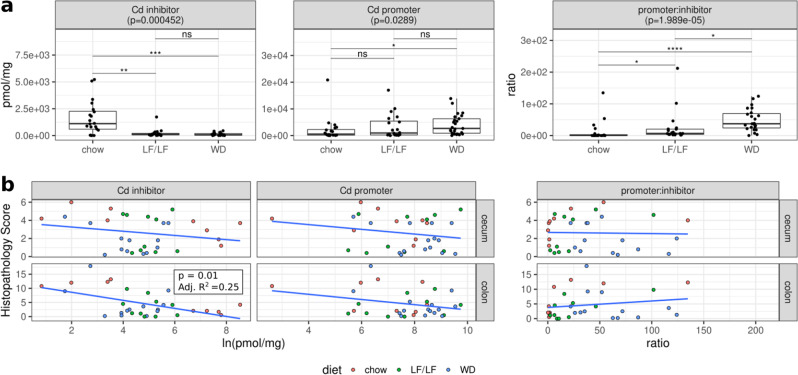
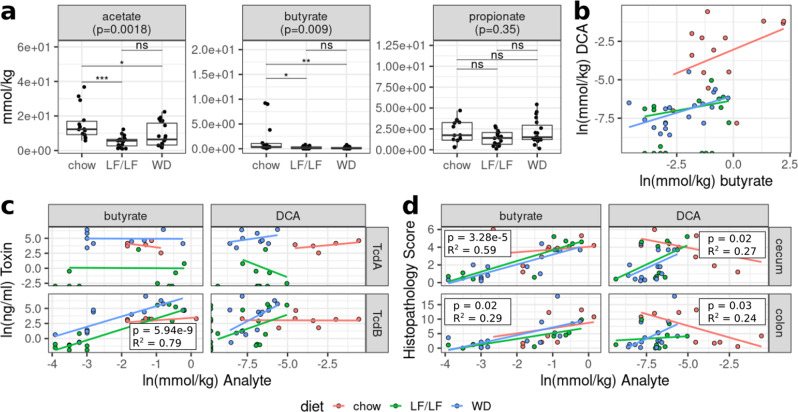
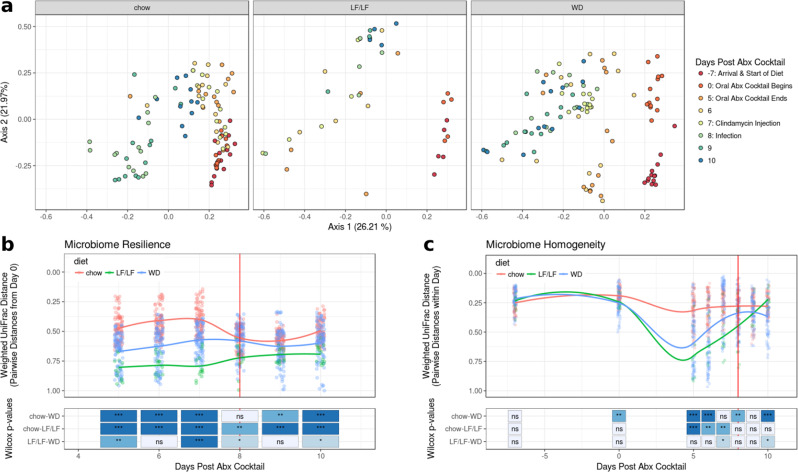
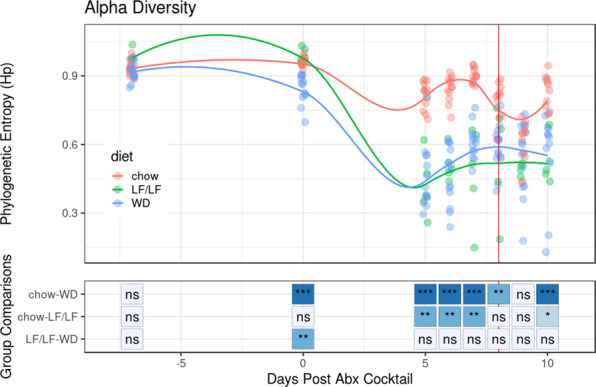
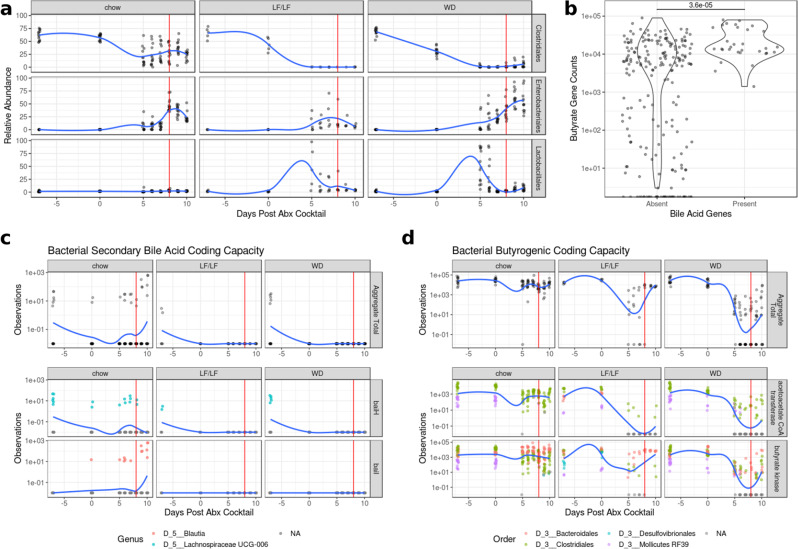
Clostridioides difficile infection (CDI) is the leading cause of hospital-acquired diarrhea, and emerging evidence has linked dietary components with CDI pathogenesis, suggesting that dietary modulation may be an effective strategy for prevention. Here, we show that mice fed a high-fat/low-fiber "Western-type" diet (WD) had dramatically increased mortality in a murine model of antibiotic-induced CDI compared to a low-fat/low-fiber (LF/LF) diet and standard mouse chow controls. We found that the WD had a pro- C. difficile bile acid composition that was driven in part by higher levels of primary bile acids that are produced to digest fat, and a lower level of secondary bile acids that are produced by the gut microbiome. This lack of secondary bile acids was associated with a greater disturbance to the gut microbiome with antibiotics in both the WD and LF/LF diet compared to mouse chow. Mice fed the WD also had the highest level of toxin TcdA just prior to the onset of mortality, but not of TcdB or increased inflammation. These findings indicate that dietary intervention to decrease fat may complement previously proposed dietary intervention strategies to prevent CDI in high-risk individuals.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
