

Pituitary Gonadotropin Gene Expression During Induced Onset of Postsmolt Maturation in Male Atlantic Salmon: In Vivo and Tissue Culture Studies
- PMID: 35370944
- PMCID: PMC8964956
- DOI: 10.3389/fendo.2022.826920
Pituitary Gonadotropin Gene Expression During Induced Onset of Postsmolt Maturation in Male Atlantic Salmon: In Vivo and Tissue Culture Studies
Abstract
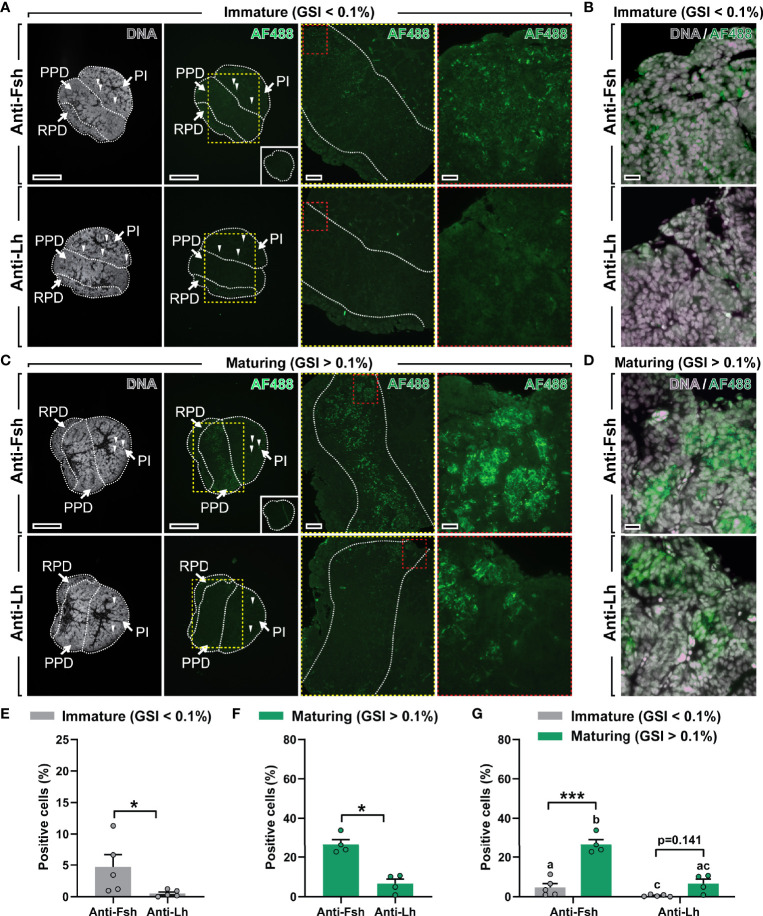
Precocious male maturation causes reduced welfare and increased production costs in Atlantic salmon (Salmo salar) aquaculture. The pituitary produces and releases follicle-stimulating hormone (Fsh), the gonadotropin triggering puberty in male salmonids. However, little is known about how Fsh production is regulated in Atlantic salmon. We examined, in vivo and ex vivo, transcriptional changes of gonadotropin-related genes accompanying the initial steps of testis maturation, in pituitaries of males exposed to photoperiod and temperature conditions promoting maturation (constant light and 16°C). Pituitary fshb, lhb and gnrhr2bba transcripts increased in vivo in maturing males (gonado-somatic index > 0.1%). RNA sequencing (RNAseq) analysis using pituitaries from genetically similar males carrying the same genetic predisposition to mature, but differing by responding or not responding to stimulatory environmental conditions, revealed 144 differentially expressed genes, ~2/3rds being up-regulated in responders, including fshb and other pituitary hormones, steroid-related and other puberty-associated transcripts. Functional enrichment analyses confirmed gene involvement in hormone/steroid production and gonad development. In ex vivo studies, whole pituitaries were exposed to a selection of hormones and growth factors. Gonadotropin-releasing hormone (Gnrh), 17β-estradiol (E2) and 11-ketotestosterone (11-KT) up-regulated gnrhr2bba and lhb, while fshb was up-regulated by Gnrh but down-regulated by 11-KT in pituitaries from immature males. Also pituitaries from maturing males responded to Gnrh and sex steroids by increased gnrhr2bba and lhb transcript levels, but fshb expression remained unchanged. Growth factors (inhibin A, activin A and insulin-like growth factor 1) did not change gnrhr2bba, lhb or fshb transcript levels in pituitaries either from immature or maturing males. Additional pituitary ex vivo studies on candidates identified by RNAseq showed that these transcripts were preferentially regulated by Gnrh and sex steroids, but not by growth factors, and that Gnrh/sex steroids were less effective when incubating pituitaries from maturing males. Our results suggest that a yet to be characterized mechanism up-regulating fshb expression in the salmon pituitary is activated in response to stimulatory environmental conditions prior to morphological signs of testis maturation, and that the transcriptional program associated with this mechanism becomes unresponsive or less responsive to most stimulators ex vivo once males had entered pubertal developmental in vivo.
Keywords: Atlantic salmon; follicle-stimulating hormone; pituitary; puberty; transcriptomics.
Copyright © 2022 Crespo, Skaftnesmo, Kjærner-Semb, Yilmaz, Norberg, Olausson, Vogelsang, Bogerd, Kleppe, Edvardsen, Andersson, Wargelius, Hansen, Fjelldal and Schulz.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






Similar articles
-
Gnrh receptor gnrhr2bbα is expressed exclusively in lhb-expressing cells in Atlantic salmon male parr.Gen Comp Endocrinol. 2020 Jan 1;285:113293. doi: 10.1016/j.ygcen.2019.113293. Epub 2019 Sep 30. Gen Comp Endocrinol. 2020. PMID: 31580881
-
Entry into puberty is reflected in changes in hormone production but not in testicular receptor expression in Atlantic salmon (Salmo salar).Reprod Biol Endocrinol. 2019 Jun 21;17(1):48. doi: 10.1186/s12958-019-0493-8. Reprod Biol Endocrinol. 2019. PMID: 31226998 Free PMC article.
-
Environmental impacts on the gonadotropic system in female Atlantic salmon (Salmo salar) during vitellogenesis: Photothermal effects on pituitary gonadotropins, ovarian gonadotropin receptor expression, plasma sex steroids and oocyte growth.Gen Comp Endocrinol. 2015 Sep 15;221:86-93. doi: 10.1016/j.ygcen.2015.02.008. Epub 2015 Feb 21. Gen Comp Endocrinol. 2015. PMID: 25712829
-
Regulation of gonadotropin subunit genes in tilapia.Comp Biochem Physiol B Biochem Mol Biol. 2001 Jun;129(2-3):489-502. doi: 10.1016/s1096-4959(01)00345-1. Comp Biochem Physiol B Biochem Mol Biol. 2001. PMID: 11399484 Review.
-
Endocrine regulation of gonadotropin and growth hormone gene transcription in fish.Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1998 Jun;119(3):325-38. doi: 10.1016/s0742-8413(98)00021-8. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 1998. PMID: 9827005 Review.
Cited by
-
Hippo-vgll3 signaling may contribute to sex differences in Atlantic salmon maturation age via contrasting adipose dynamics.Biol Sex Differ. 2025 Apr 2;16(1):23. doi: 10.1186/s13293-025-00705-8. Biol Sex Differ. 2025. PMID: 40176157 Free PMC article.
-
Loss of Fshr Prevents Testicular Maturation in Atlantic Salmon (Salmo salar L.).Endocrinology. 2024 Feb 20;165(4):bqae013. doi: 10.1210/endocr/bqae013. Endocrinology. 2024. PMID: 38298132 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources