

Genome Engineering Technology for Durable Disease Resistance: Recent Progress and Future Outlooks for Sustainable Agriculture
- PMID: 35371164
- PMCID: PMC8968944
- DOI: 10.3389/fpls.2022.860281
Genome Engineering Technology for Durable Disease Resistance: Recent Progress and Future Outlooks for Sustainable Agriculture
Abstract
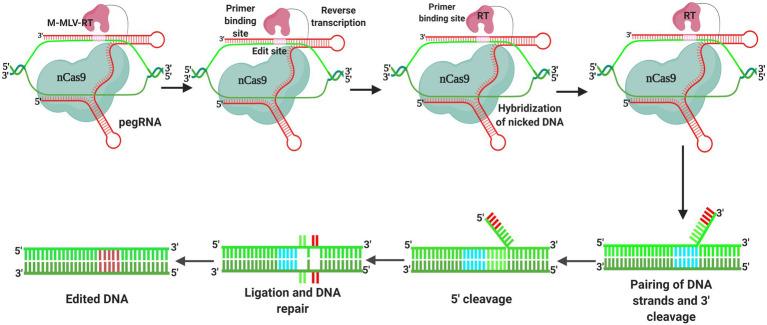
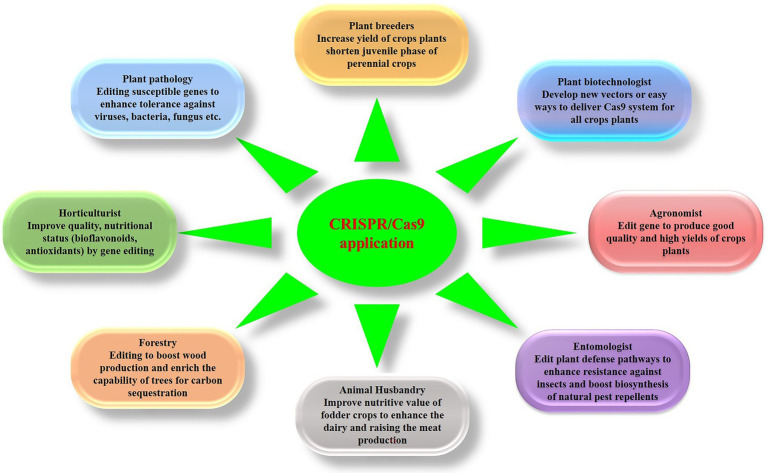
Crop production worldwide is under pressure from multiple factors, including reductions in available arable land and sources of water, along with the emergence of new pathogens and development of resistance in pre-existing pathogens. In addition, the ever-growing world population has increased the demand for food, which is predicted to increase by more than 100% by 2050. To meet these needs, different techniques have been deployed to produce new cultivars with novel heritable mutations. Although traditional breeding continues to play a vital role in crop improvement, it typically involves long and laborious artificial planting over multiple generations. Recently, the application of innovative genome engineering techniques, particularly CRISPR-Cas9-based systems, has opened up new avenues that offer the prospects of sustainable farming in the modern agricultural industry. In addition, the emergence of novel editing systems has enabled the development of transgene-free non-genetically modified plants, which represent a suitable option for improving desired traits in a range of crop plants. To date, a number of disease-resistant crops have been produced using gene-editing tools, which can make a significant contribution to overcoming disease-related problems. Not only does this directly minimize yield losses but also reduces the reliance on pesticide application, thereby enhancing crop productivity that can meet the globally increasing demand for food. In this review, we describe recent progress in genome engineering techniques, particularly CRISPR-Cas9 systems, in development of disease-resistant crop plants. In addition, we describe the role of CRISPR-Cas9-mediated genome editing in sustainable agriculture.
Keywords: CRISPR-Cas system; disease resistance; genome editing; pesticide; plant pathogen.
Copyright © 2022 Ali, Yu, Hussain, Ali, Ahmar, Sohail, Riaz, Ashraf, Abdalmegeed, Wang, Imran, Manghwar and Zhou.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
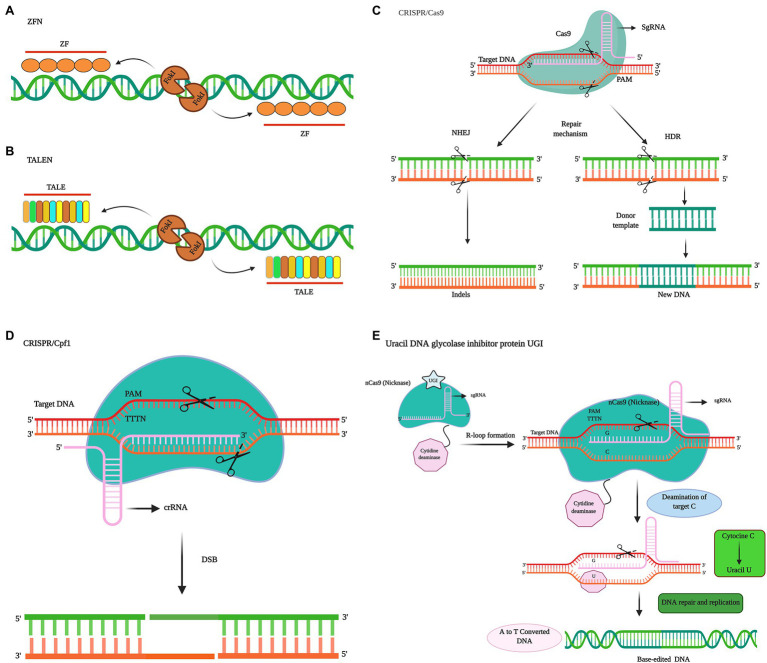
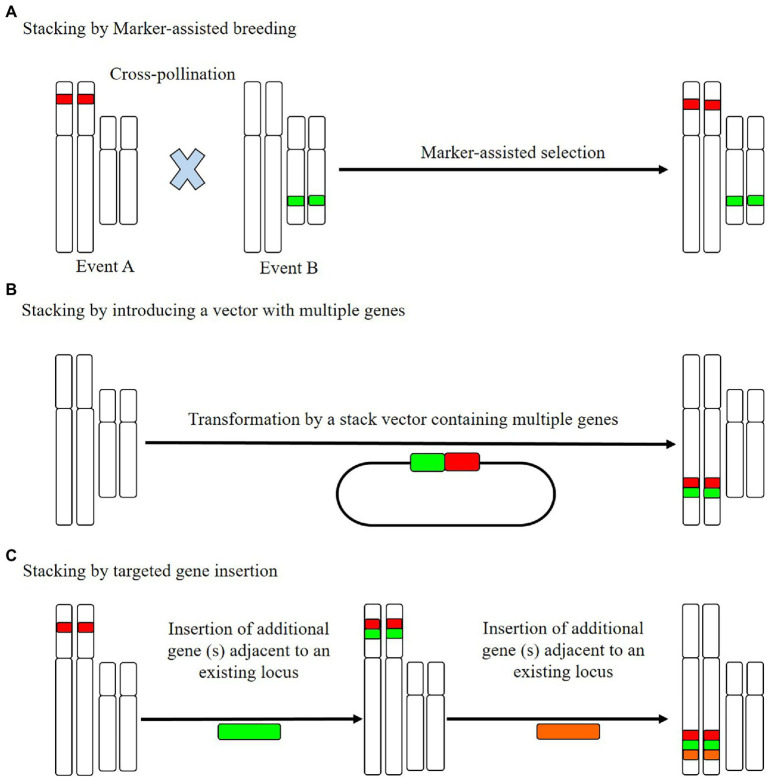
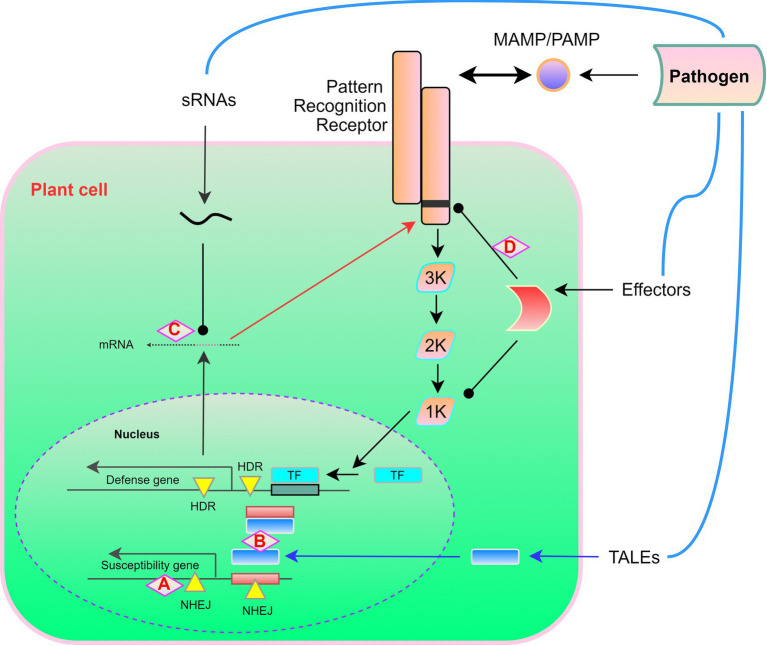
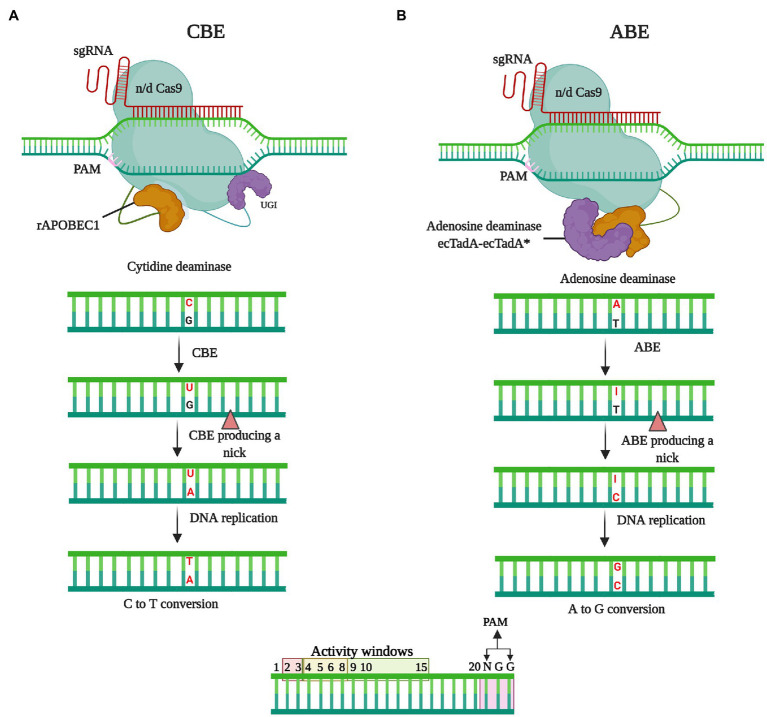
References
-
- Acevedo-Garcia J., Spencer D., Thieron H., Reinstädler A., Hammond-Kosack K., Phillips A. L., et al. (2017). mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach. Plant Biotechnol. J. 15, 367–378. doi: 10.1111/pbi.12631, PMID: - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources