

Disease-associated KBTBD4 mutations in medulloblastoma elicit neomorphic ubiquitylation activity to promote CoREST degradation
- PMID: 35379950
- PMCID: PMC9525703
- DOI: 10.1038/s41418-022-00983-4
Disease-associated KBTBD4 mutations in medulloblastoma elicit neomorphic ubiquitylation activity to promote CoREST degradation
Abstract
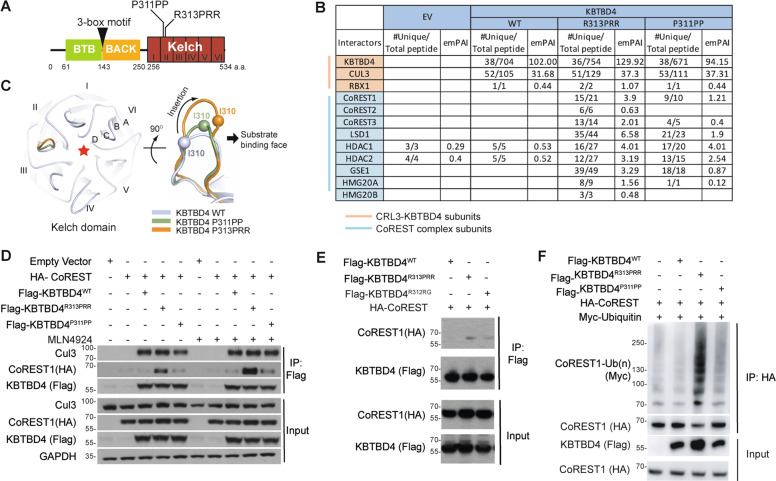
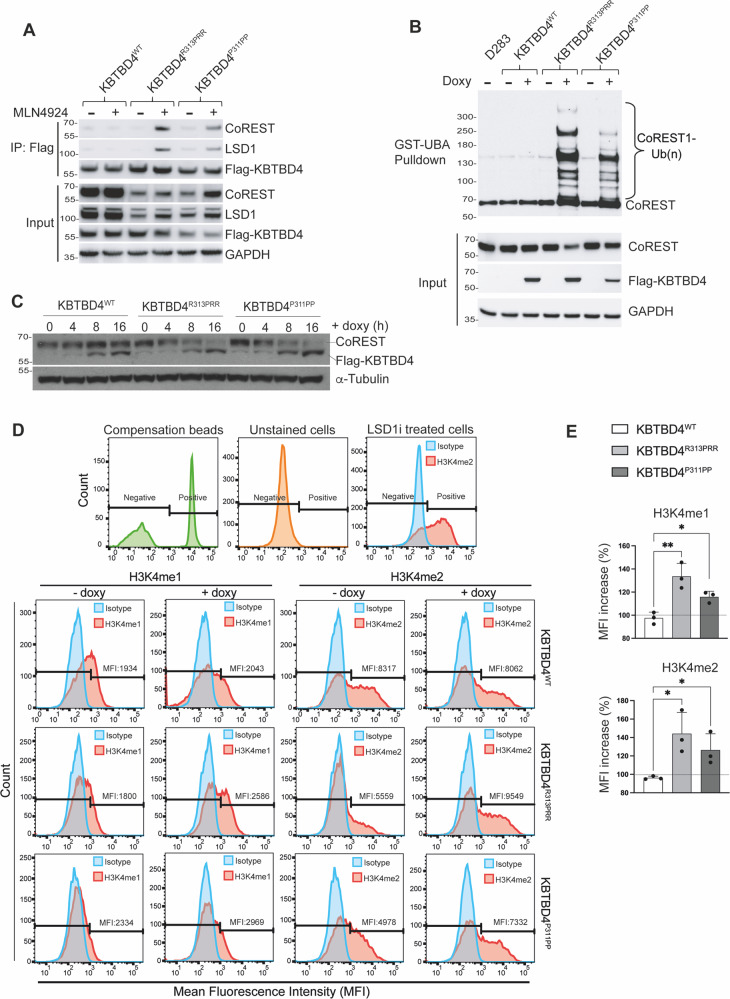
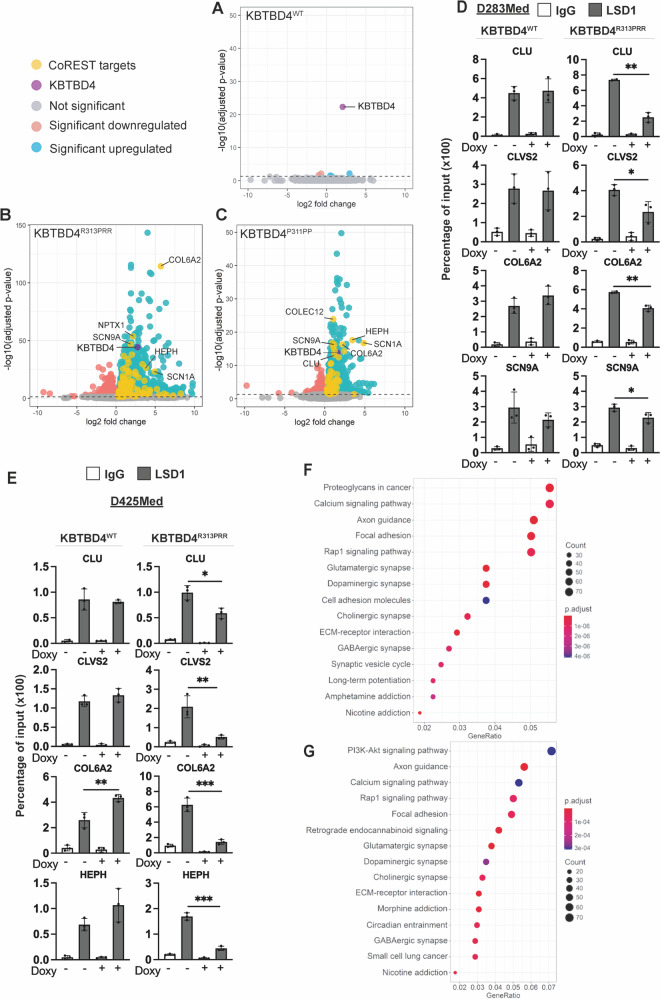
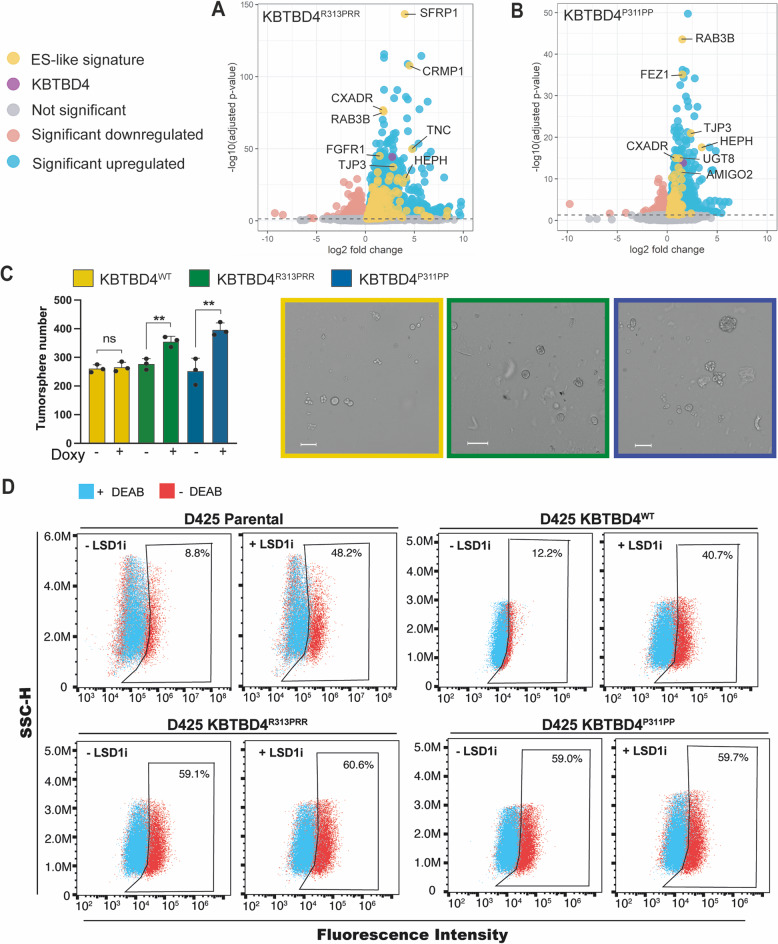
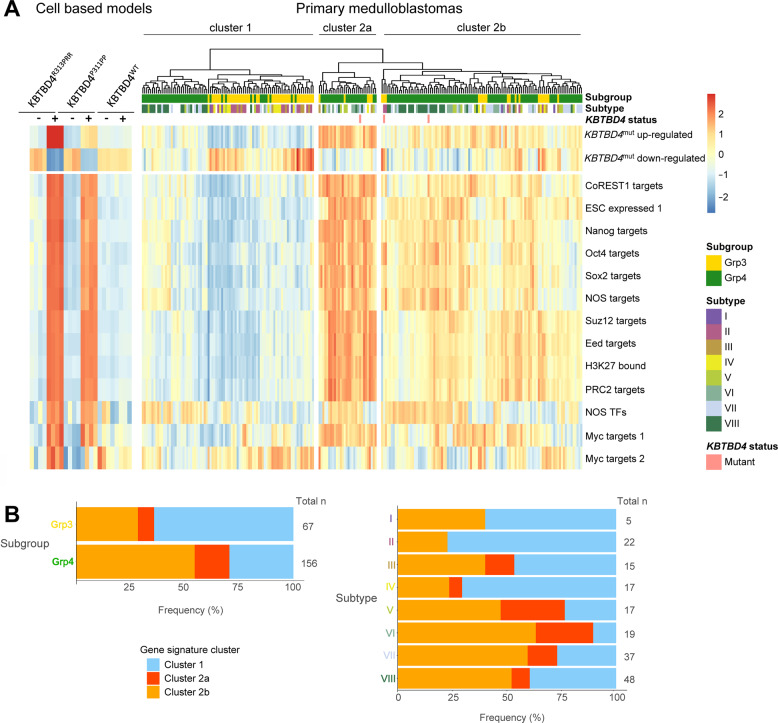
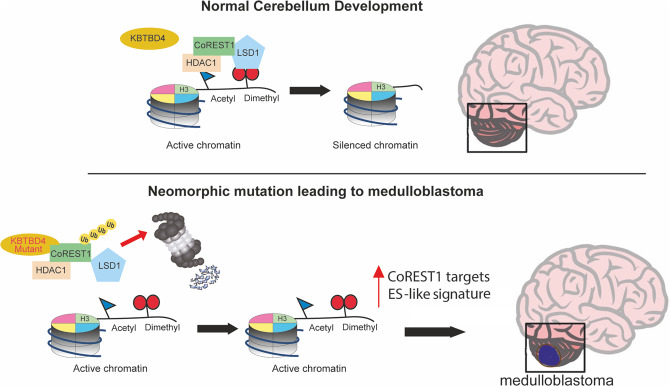
Medulloblastoma is the most common malignant brain tumour in children. Genomic studies have identified distinct disease subgroups: wnt/wingless (WNT), sonic hedgehog (SHH), and non-WNT/non-SHH, comprising group 3 and group 4. Alterations in WNT and SHH signalling form the pathogenetic basis for their subgroups, whereas those for non-WNT/non-SHH tumours remain largely elusive. Recent analyses have revealed recurrent in-frame insertions in the E3 ubiquitin ligase adaptor Kelch Repeat and BTB Domain Containing 4 (KBTBD4) in cases of group 3/4 medulloblastoma. Critically, group 3/4 tumours with KBTBD4 mutations typically lack other gene-specific alterations, such as MYC amplification, indicating KBTBD4 insertion mutations as the primary genetic driver. Delineating the role of KBTBD4 mutations thus offers significant opportunities to understand tumour pathogenesis and to exploit the underpinning mechanisms therapeutically. Here, we show a novel mechanism in cancer pathogenesis whereby indel mutations in KBTBD4 drive its recognition of neo-substrates for degradation. We observe that KBTBD4 mutants promote the recruitment and ubiquitylation of the REST Corepressor (CoREST), which forms a complex to modulate chromatin accessibility and transcriptional programmes. The degradation of CoREST promoted by KBTBD4 mutation diverts epigenetic programmes inducing significant alterations in transcription to promote increased stemness of cancer cells. Transcriptional analysis of >200 human group 3 and 4 medulloblastomas by RNA-seq, highlights the presence of CoREST and stem-like signatures in tumours with KBTBD4 mutations, which extend to a further sub-set of non-mutant tumours, suggesting CoREST alterations as a novel pathogenetic mechanism of wide relevance in groups 3 and 4. Our findings uncover KBTBD4 mutation as a novel driver of epigenetic reprogramming in non-WNT/non-SHH medulloblastoma, establish a novel mode of tumorigenesis through gain-of-function mutations in ubiquitin ligases (neo-substrate recruitment) and identify both mutant KBTBD4 and CoREST complexes as new druggable targets for improved tumour-specific therapies.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
