

Apolar chemical environments compact unfolded RNAs and can promote folding
- PMID: 35382036
- PMCID: PMC8978554
- DOI: 10.1016/j.bpr.2021.100004
Apolar chemical environments compact unfolded RNAs and can promote folding
Abstract
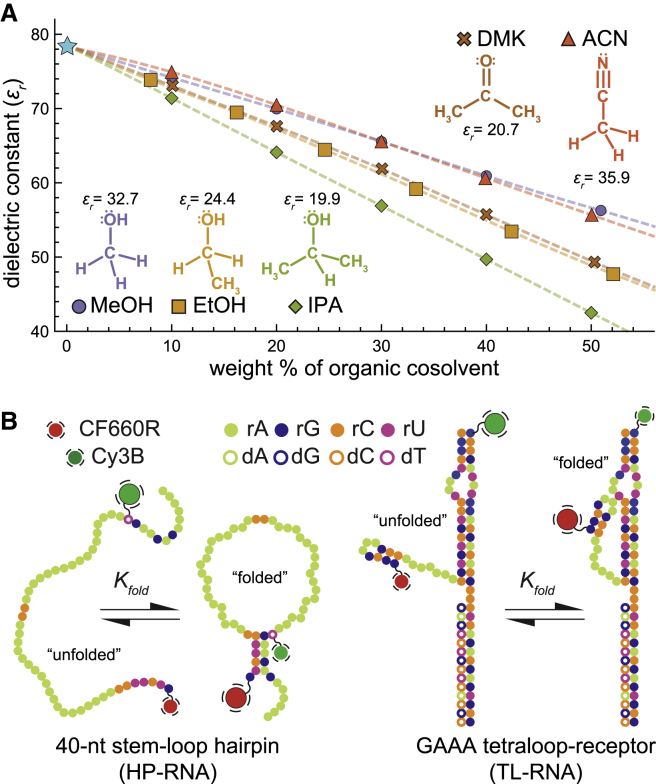
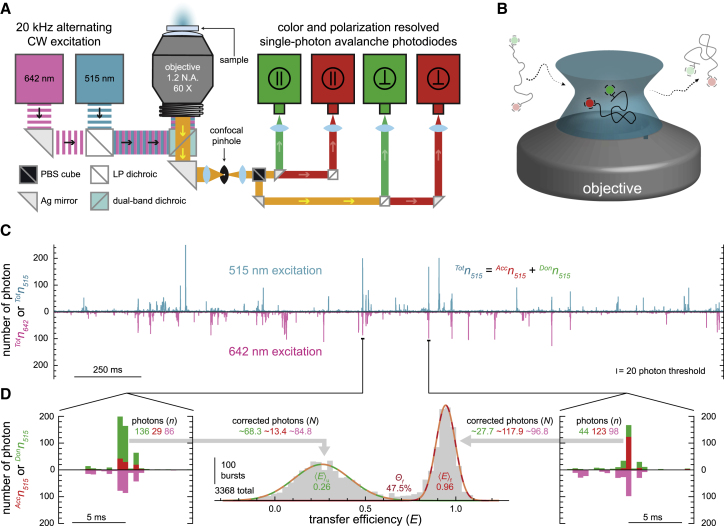
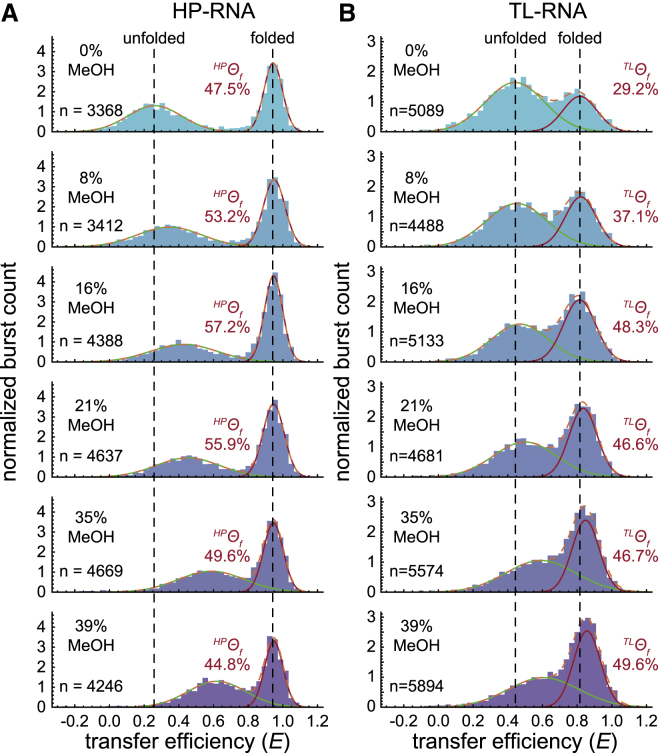
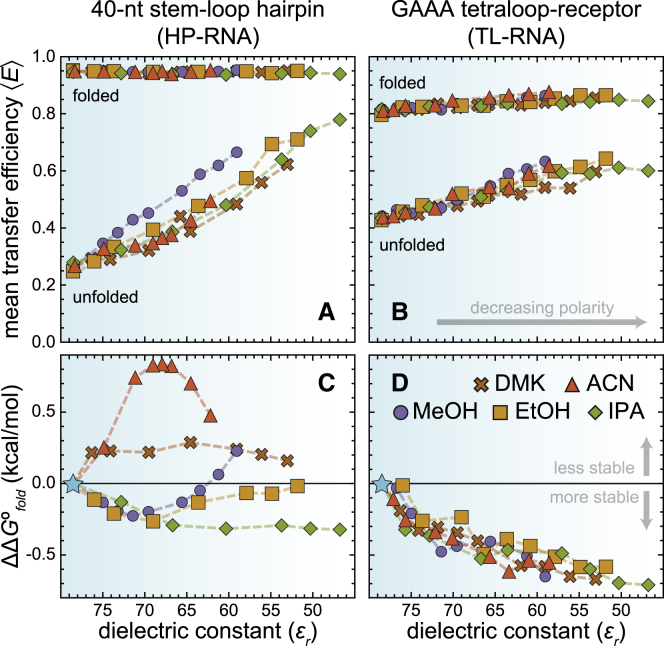
It is well documented that the structure, and thus function, of nucleic acids depends on the chemical environment surrounding them, which often includes potential proteinaceous binding partners. The nonpolar amino acid side chains of these proteins will invariably alter the polarity of the local chemical environment around the nucleic acid. However, we are only beginning to understand how environmental polarity generally influences the structural and energetic properties of RNA folding. Here, we use a series of aqueous-organic cosolvent mixtures to systematically modulate the solvent polarity around two different RNA folding constructs that can form either secondary or tertiary structural elements. Using single-molecule Förster resonance energy transfer spectroscopy to simultaneously monitor the structural and energetic properties of these RNAs, we show that the unfolded conformations of both model RNAs become more compact in apolar environments characterized by dielectric constants less than that of pure water. In the case of tertiary structure formation, this compaction also gives rise to more energetically favorable folding. We propose that these physical changes arise from an enhanced accumulation of counterions in the low dielectric environment surrounding the unfolded RNA.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Kolev S.K., Petkov P.S., et al. Vayssilov G.N. Interaction of Na+, K+, Mg2+ and Ca2+ counter cations with RNA. Metallomics. 2018;10:659–678. - PubMed
-
- Fiore J.L., Holmstrom E.D., et al. Nesbitt D.J. The role of counterion valence and size in GAAA tetraloop-receptor docking/undocking kinetics. J. Mol. Biol. 2012;423:198–216. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
