

The role of activation of two different sGC binding sites by NO-dependent and NO-independent mechanisms in the regulation of SACs in rat ventricular cardiomyocytes
- PMID: 35384354
- PMCID: PMC8981922
- DOI: 10.14814/phy2.15246
The role of activation of two different sGC binding sites by NO-dependent and NO-independent mechanisms in the regulation of SACs in rat ventricular cardiomyocytes
Abstract
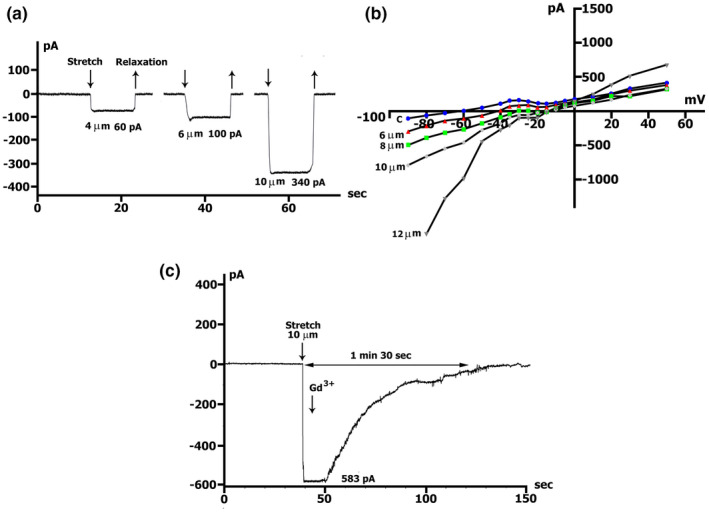
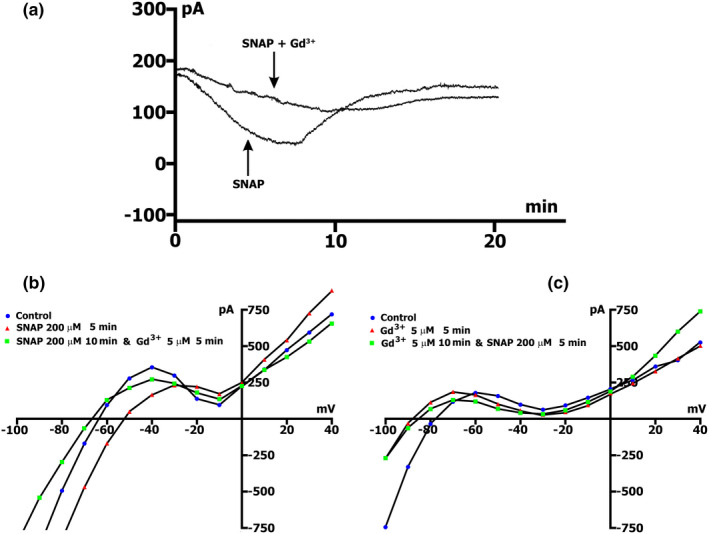
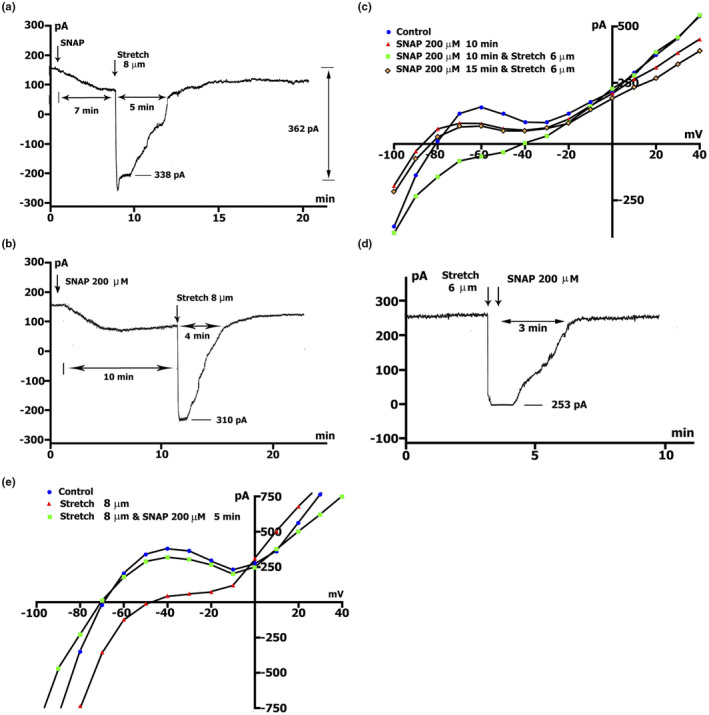
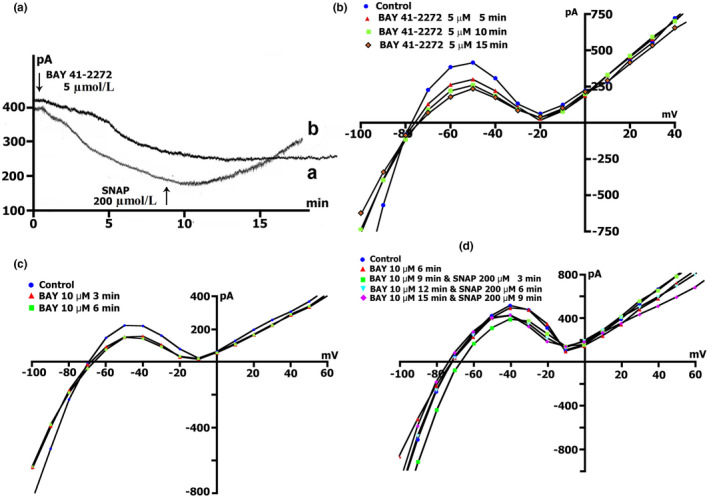
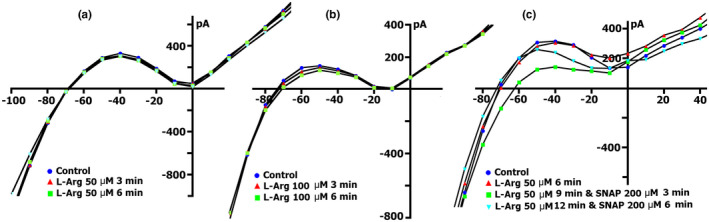
The mechanoelectrical feedback (MEF) mechanism in the heart that plays a significant role in the occurrence of arrhythmias, involves cation flux through cation nonselective stretch-activated channels (SACs). It is well known that nitric oxide (NO) can act as a regulator of MEF. Here we addressed the possibility of SAC's regulation along NO-dependent and NO-independent pathways, as well as the possibility of S-nitrosylation of SACs. In freshly isolated rat ventricular cardiomyocytes, using the patch-clamp method in whole-cell configuration, inward nonselective stretch-activated cation current ISAC was recorded through SACs, which occurs during dosed cell stretching. NO donor SNAP, α1-subunit of sGC activator BAY41-2272, sGC blocker ODQ, PKG blocker KT5823, PKG activator 8Br-cGMP, and S-nitrosylation blocker ascorbic acid, were employed. We concluded that the physiological concentration of NO in the cell is a necessary condition for the functioning of SACs. An increase in NO due to SNAP in an unstretched cell causes the appearance of a Gd3+ -sensitive nonselective cation current, an analog of ISAC , while in a stretched cell it eliminates ISAC . The NO-independent pathway of sGC activation of α subunit, triggered by BAY41-2272, is also important for the regulation of SACs. Since S-nitrosylation inhibitor completely abolishes ISAC , this mechanism occurs. The application of BAY41-2272 cannot induce ISAC in a nonstretched cell; however, the addition of SNAP on its background activates SACs, rather due to S-nitrosylation. ODQ eliminates ISAC , but SNAP added on the background of stretch increases ISAC in addition to ODQ. This may be a result of the lack of NO as a result of inhibition of NOS by metabolically modified ODQ. KT5823 reduces PKG activity and reduces SACs phosphorylation, leading to an increase in ISAC . 8Br-cGMP reduces ISAC by activating PKG and its phosphorylation. These results demonstrate a significant contribution of S-nitrosylation to the regulation of SACs.
Keywords: 8Br-cGMP; BAY41-2272; KT5823; L-Arginine; ODQ; ascorbic acid; nitric oxide; nitric oxide synthase; patch-clamp, SNAP; soluble guanylyl cyclase; stretch-activated channels; ventricular cardiomyocytes.
© 2022 The Authors. Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
None declared.
Figures










Similar articles
-
Feedback control through cGMP-dependent protein kinase contributes to differential regulation and compartmentation of cGMP in rat cardiac myocytes.Circ Res. 2010 Nov 12;107(10):1232-40. doi: 10.1161/CIRCRESAHA.110.226712. Epub 2010 Sep 16. Circ Res. 2010. PMID: 20847310
-
Nitric oxide-cGMP-protein kinase G pathway negatively regulates vascular transient receptor potential channel TRPC6.J Physiol. 2008 Sep 1;586(17):4209-23. doi: 10.1113/jphysiol.2008.156083. Epub 2008 Jul 10. J Physiol. 2008. PMID: 18617565 Free PMC article.
-
Mechanotransduction-Driven Modulation of L-Type Calcium Channels: Roles of Nitric Oxide, S-Nitrosylation, and cGMP in Rat Ventricular Cardiomyocytes.Int J Mol Sci. 2025 Aug 4;26(15):7539. doi: 10.3390/ijms26157539. Int J Mol Sci. 2025. PMID: 40806667 Free PMC article.
-
Isolated Cardiomyocytes: Mechanosensitivity of Action Potential, Membrane Current and Ion Concentration.In: Kamkin A, Kiseleva I, editors. Mechanosensitivity in Cells and Tissues. Moscow: Academia; 2005. In: Kamkin A, Kiseleva I, editors. Mechanosensitivity in Cells and Tissues. Moscow: Academia; 2005. PMID: 21290777 Free Books & Documents. Review.
-
Mechano-Electric Feedback in the Heart: Evidence from Intracellular Microelectrode Recordings on Multicellular Preparations and Single Cells from Healthy and Diseased Tissue.In: Kamkin A, Kiseleva I, editors. Mechanosensitivity in Cells and Tissues. Moscow: Academia; 2005. In: Kamkin A, Kiseleva I, editors. Mechanosensitivity in Cells and Tissues. Moscow: Academia; 2005. PMID: 21290768 Free Books & Documents. Review.
Cited by
-
Simulated Microgravity Attenuates Stretch Sensitivity of Mechanically Gated Channels in Rat Ventricular Myocytes.Int J Mol Sci. 2025 Jul 11;26(14):6653. doi: 10.3390/ijms26146653. Int J Mol Sci. 2025. PMID: 40724903 Free PMC article.
-
Simulated Microgravity and Hypergravity Affect the Expression Level of Soluble Guanylate Cyclase, Adenylate Cyclase, and Phosphodiesterase Genesin Rat Ventricular Cardiomyocytes.Bull Exp Biol Med. 2024 Jan;176(3):359-362. doi: 10.1007/s10517-024-06024-z. Epub 2024 Feb 12. Bull Exp Biol Med. 2024. PMID: 38342810
-
Transcriptomic profile of the mechanosensitive ion channelome in human cardiac fibroblasts.Exp Biol Med (Maywood). 2023 Dec;248(23):2341-2350. doi: 10.1177/15353702231218488. Epub 2023 Dec 30. Exp Biol Med (Maywood). 2023. PMID: 38158807 Free PMC article.
-
Identification of RNA reads encoding different channels in isolated rat ventricular myocytes and the effect of cell stretching on L-type Ca2+current.Biol Direct. 2023 Oct 30;18(1):70. doi: 10.1186/s13062-023-00427-0. Biol Direct. 2023. PMID: 37899484 Free PMC article.
-
Tetrahydrocurcumin-Related Vascular Protection: An Overview of the Findings from Animal Disease Models.Molecules. 2022 Aug 10;27(16):5100. doi: 10.3390/molecules27165100. Molecules. 2022. PMID: 36014335 Free PMC article. Review.
References
-
- Barouch, L. A. , Harrison, R. W. , Skaf, M. W. , Rosas, G. O. , Cappola, T. P. , Kobeissi, Z. A. , Hobai, I. A. , Lemmon, C. A. , Burnett, A. L. , & O’Rourke, B. (2002). Nitric oxide regulates the heart by spatial confinement of nitric oxide synthase isoforms. Nature, 416(6878), 337–339. - PubMed
-
- Becker, E. M. , Alonso‐Alija, C. , Apeler, H. , Gerzer, R. , Minuth, T. , Pleiss, U. , Schmidt, P. , Schramm, M. , Schröder, H. , Schroeder, W. , Steinke, W. , Straub, A. , & Stasch, J. P. (2001). NO‐independent regulatory site of direct sGC stimulators like YC‐1 and BAY 41–2272. BioMed Central Pharmacology, 1, 13. - PMC - PubMed
-
- Belus, A. , & White, E. (2002). Effects of streptomycin sulfate on I CaL , I Kr, and I Ks in guinea‐pig ventricular myocytes. European Journal of Pharmacology, 445(3), 171–178. - PubMed
-
- Boycott, H. E. , Barbier, C. S. M. , Eichel, C. A. , Costa, K. D. , Martins, R. P. , Louault, F. , Dilanian, G. , Coulombe, A. , Hatem, S. N. , & Balse, E. (2013). Shear stress triggers insertion of voltage‐gated potassium channels from intracellular compartments in atrial myocytes. Proceedings of the National Academy of Sciences USA, 110(41), E3955–E3964. 10.1073/pnas.1309896110 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
