

Control of coronary vascular resistance by eicosanoids via a novel GPCR
- PMID: 35385329
- PMCID: PMC9255704
- DOI: 10.1152/ajpcell.00454.2021
Control of coronary vascular resistance by eicosanoids via a novel GPCR
Erratum in
-
Corrigendum for Alkayed et al., volume 322, 2022, p. C1011-C1021.Am J Physiol Cell Physiol. 2022 Jun 1;322(6):C1270. doi: 10.1152/ajpcell.00454.2021_cor. Am J Physiol Cell Physiol. 2022. PMID: 35658714 Free PMC article. No abstract available.
Abstract
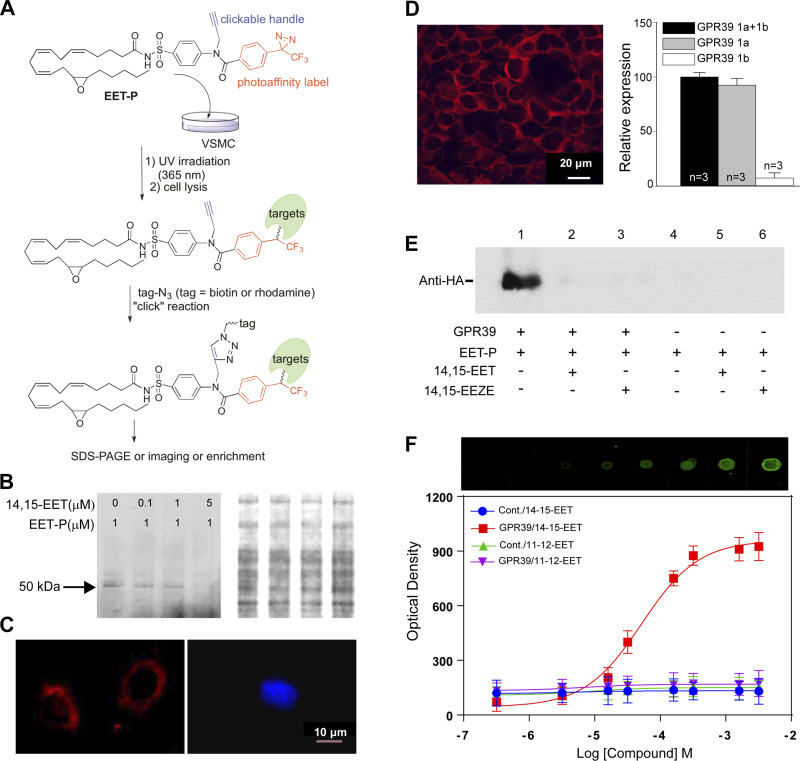
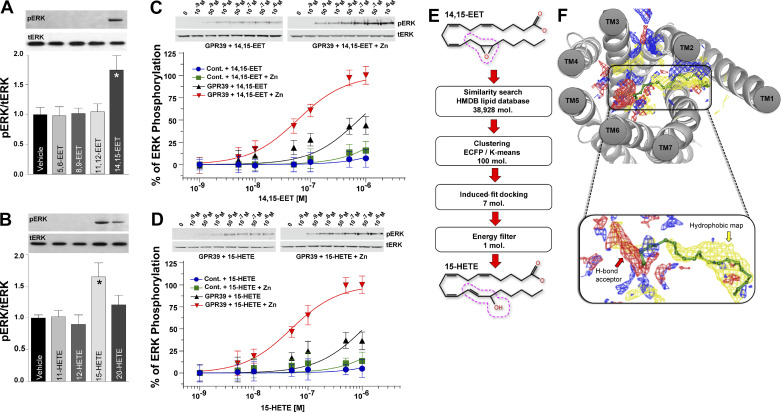
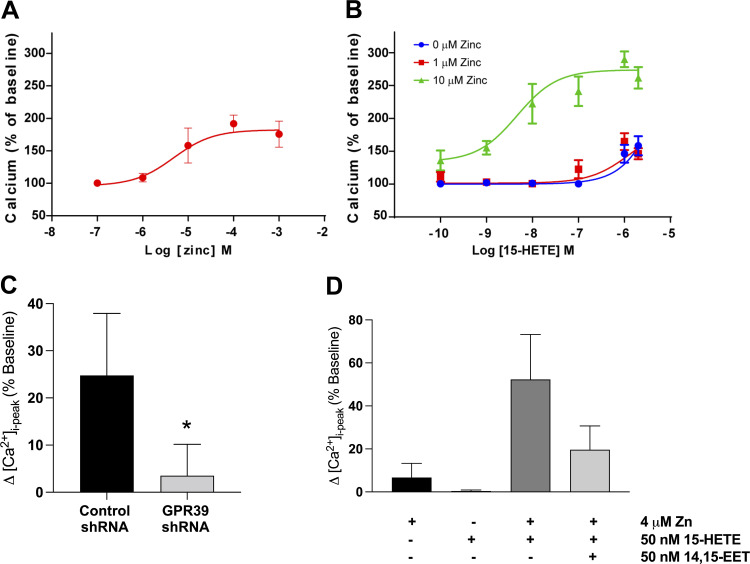
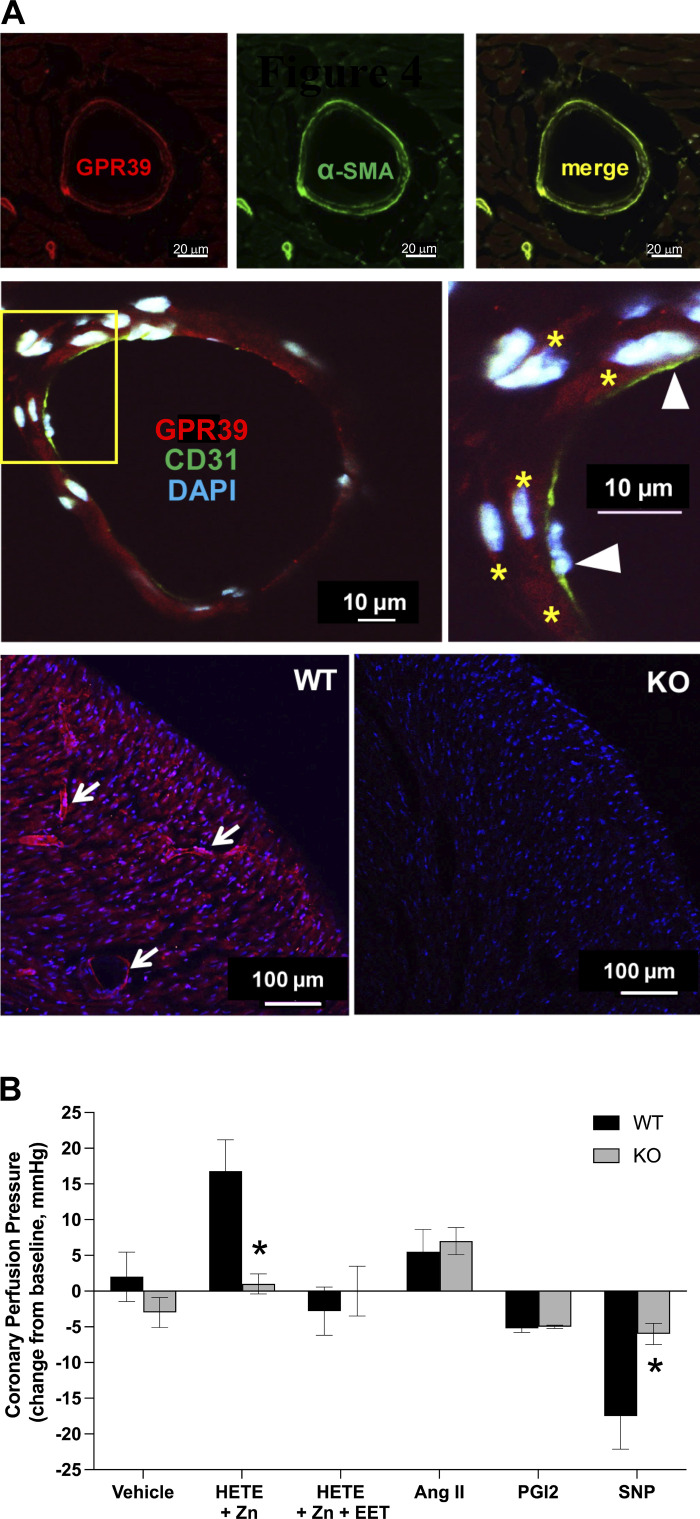
Arachidonic acid metabolites epoxyeicosatrienoates (EETs) and hydroxyeicosatetraenoates (HETEs) are important regulators of myocardial blood flow and coronary vascular resistance (CVR), but their mechanisms of action are not fully understood. We applied a chemoproteomics strategy using a clickable photoaffinity probe to identify G protein-coupled receptor 39 (GPR39) as a microvascular smooth muscle cell (mVSMC) receptor selective for two endogenous eicosanoids, 15-HETE and 14,15-EET, which act on the receptor to oppose each other's activity. The former increases mVSMC intracellular calcium via GPR39 and augments coronary microvascular resistance, and the latter inhibits these actions. Furthermore, we find that the efficacy of both ligands is potentiated by zinc acting as an allosteric modulator. Measurements of coronary perfusion pressure (CPP) in GPR39-null hearts using the Langendorff preparation indicate the receptor senses these eicosanoids to regulate microvascular tone. These results implicate GPR39 as an eicosanoid receptor and key regulator of myocardial tissue perfusion. Our findings will have a major impact on understanding the roles of eicosanoids in cardiovascular physiology and disease and provide an opportunity for the development of novel GPR39-targeting therapies for cardiovascular disease.
Keywords: EETs; GPCR; GPR39; HETEs; eicosanoids.
Conflict of interest statement
This research involves technology of which N. J. Alkayed, S. Kaul, and S. Nagarajan are co-inventors and which has been licensed, in part by OHSU, to Vasocardea. OHSU and S. Kaul have a financial interest in Vasocardea, a company that may have a commercial interest in the results of this research and technology. This potential conflict of interest has been reviewed and managed by Oregon Health and Science University. None of the other authors has any conflicts of interest, financial or otherwise, to disclose.
This article is part of the special collection "Advances in GPCRs: Structure, Mechanisms, Disease, and Pharmacology." Wei Kong, MD, PhD, and Jinpeng Sun, PhD, served as Guest Editors of this collection.
Figures
References
-
- Muller JM, Davis MJ, Chilian WM. Integrated regulation of pressure and flow in the coronary microcirculation. Cardiovasc Res 32: 668–678, 1996. - PubMed
-
- Capdevila JH, Falck JR, Harris RC. Cytochrome P450 and arachidonic acid bioactivation. Molecular and functional properties of the arachidonate monooxygenase. J Lipid Res 41: 163–181, 2000. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases