

Mutational background influences P. aeruginosa ciprofloxacin resistance evolution but preserves collateral sensitivity robustness
- PMID: 35385351
- PMCID: PMC9169633
- DOI: 10.1073/pnas.2109370119
Mutational background influences P. aeruginosa ciprofloxacin resistance evolution but preserves collateral sensitivity robustness
Abstract
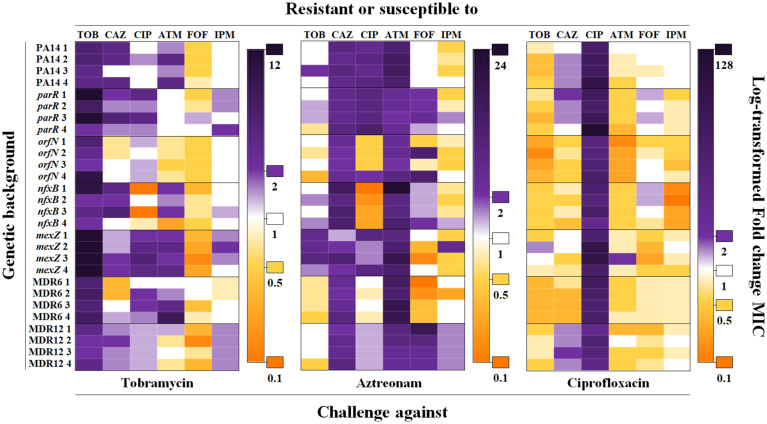
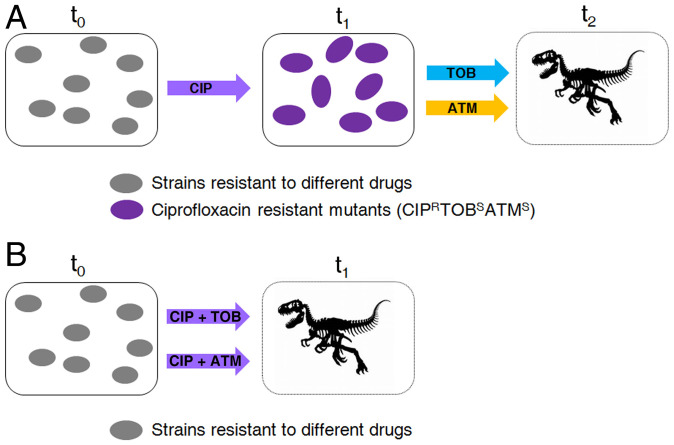
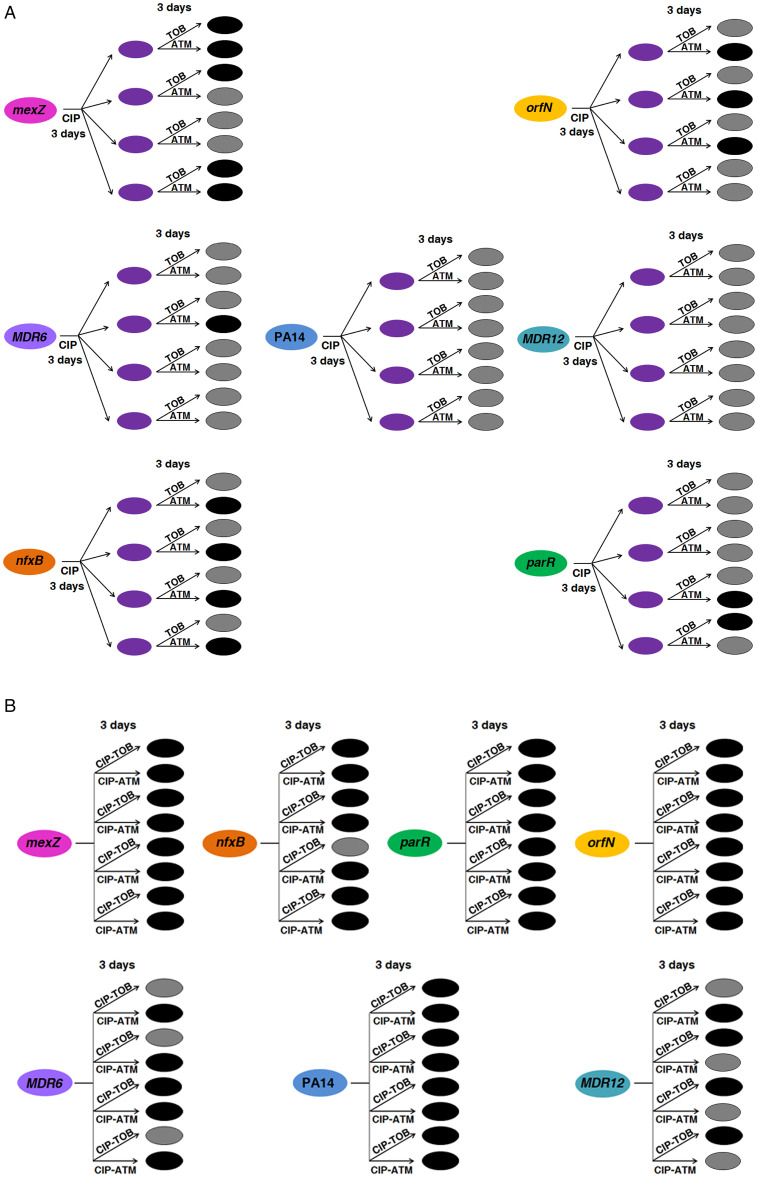
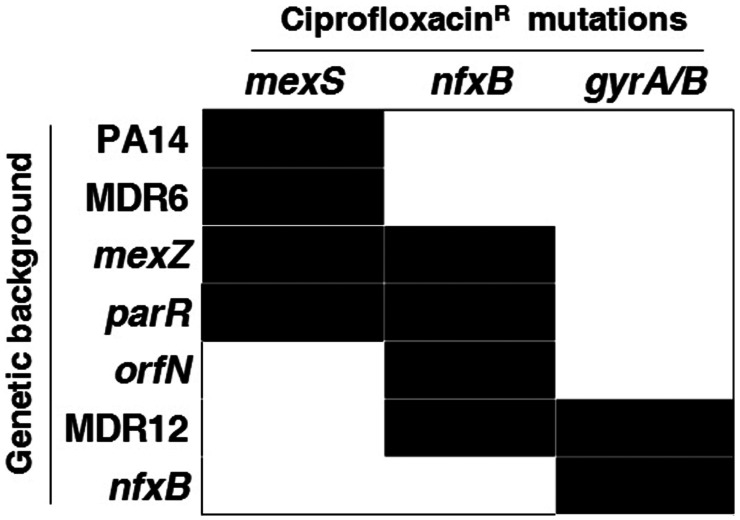
Collateral sensitivity is an evolutionary trade-off whereby acquisition of the adaptive phenotype of resistance to an antibiotic leads to the nonadaptive increased susceptibility to another. The feasibility of harnessing such a trade-off to design evolutionary-based approaches for treating bacterial infections has been studied using model strains. However, clinical application of collateral sensitivity requires its conservation among strains presenting different mutational backgrounds. Particularly relevant is studying collateral sensitivity robustness of already-antibiotic-resistant mutants when challenged with a new antimicrobial, a common situation in clinics that has hardly been addressed. We submitted a set of diverse Pseudomonas aeruginosa antibiotic-resistant mutants to short-term evolution in the presence of different antimicrobials. Ciprofloxacin selects different clinically relevant resistance mutations in the preexisting resistant mutants, which gave rise to the same, robust, collateral sensitivity to aztreonam and tobramycin. We then experimentally determined that alternation of ciprofloxacin with aztreonam is more efficient than ciprofloxacin–tobramycin alternation in driving the extinction of the analyzed antibiotic-resistant mutants. Also, we show that the combinations ciprofloxacin–aztreonam or ciprofloxacin–tobramycin are the most effective strategies for eliminating the tested P. aeruginosa antibiotic-resistant mutants. These findings support that the identification of conserved collateral sensitivity patterns may guide the design of evolution-based strategies to treat bacterial infections, including those due to antibiotic-resistant mutants. Besides, this is an example of phenotypic convergence in the absence of parallel evolution that, beyond the antibiotic-resistance field, could facilitate the understanding of evolution processes, where the selective forces giving rise to new, not clearly adaptive phenotypes remain unclear.
Keywords: Pseudomonas aeruginosa; antibiotic resistance; collateral sensitivity; convergent evolution; phenotypic convergence.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Andersson D. I., et al. , Antibiotic resistance: Turning evolutionary principles into clinical reality. FEMS Microbiol. Rev. 44, 171–188 (2020). - PubMed
-
- Laxminarayan R., Antibiotic effectiveness: Balancing conservation against innovation. Science 345, 1299–1301 (2014). - PubMed
-
- Imamovic L., Sommer M. O., Use of collateral sensitivity networks to design drug cycling protocols that avoid resistance development. Sci. Transl. Med. 5, 204ra132 (2013). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
