

Dynamic Changes in the Proteome of Early Bovine Embryos Developed In Vivo
- PMID: 35386205
- PMCID: PMC8979002
- DOI: 10.3389/fcell.2022.863700
Dynamic Changes in the Proteome of Early Bovine Embryos Developed In Vivo
Abstract
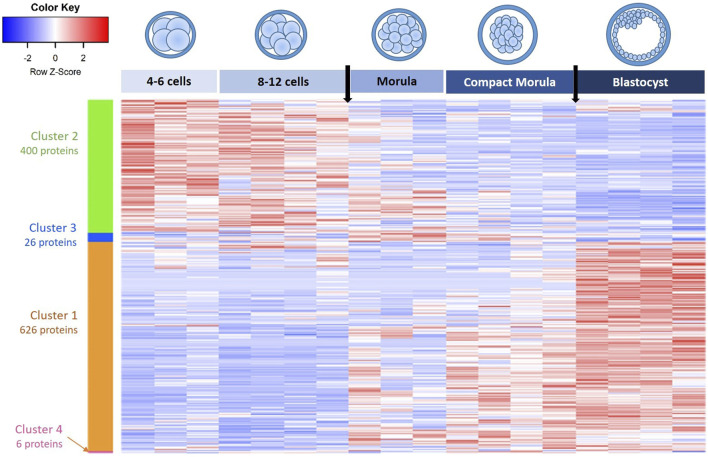
Early embryo development is a dynamic process involving important molecular and structural changes leading to the embryonic genome activation (EGA) and early cell lineage differentiation. Our aim was to elucidate proteomic changes in bovine embryos developed in vivo. Eleven females were used as embryo donors and pools of embryos at the 4-6 cell, 8-12 cell, morula, compact morula and blastocyst stages were analyzed by nanoliquid chromatography coupled with label free quantitative mass spectrometry. A total of 2,757 proteins were identified, of which 1,950 were quantitatively analyzed. Principal component analysis of data showed a clear separation of embryo pools according to their developmental stage. The hierarchical clustering of differentially abundant proteins evidenced a first cluster of 626 proteins that increased in abundance during development and a second cluster of 400 proteins that decreased in abundance during development, with most significant changes at the time of EGA and blastocyst formation. The main pathways and processes overrepresented among upregulated proteins were RNA metabolism, protein translation and ribosome biogenesis, whereas Golgi vesicle transport and protein processing in endoplasmic reticulum were overrepresented among downregulated proteins. The pairwise comparison between stages allowed us to identify specific protein interaction networks and metabolic pathways at the time of EGA, morula compaction and blastocyst formation. This is the first comprehensive study of proteome dynamics in non-rodent mammalian embryos developed in vivo. These data provide a number of protein candidates that will be useful for further mechanistic and functional studies.
Keywords: blastocyst; cattle; development; embryo; mass spectrometry; morula; proteomics.
Copyright © 2022 Banliat, Mahé, Lavigne, Com, Pineau, Labas, Guyonnet, Mermillod and Saint-Dizier.
Conflict of interest statement
Authors CB and BG were employed by the company Union Evolution. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures









References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases