

Salience by competitive and recurrent interactions: Bridging neural spiking and computation in visual attention
- PMID: 35389715
- PMCID: PMC10365548
- DOI: 10.1037/rev0000366
Salience by competitive and recurrent interactions: Bridging neural spiking and computation in visual attention
Abstract
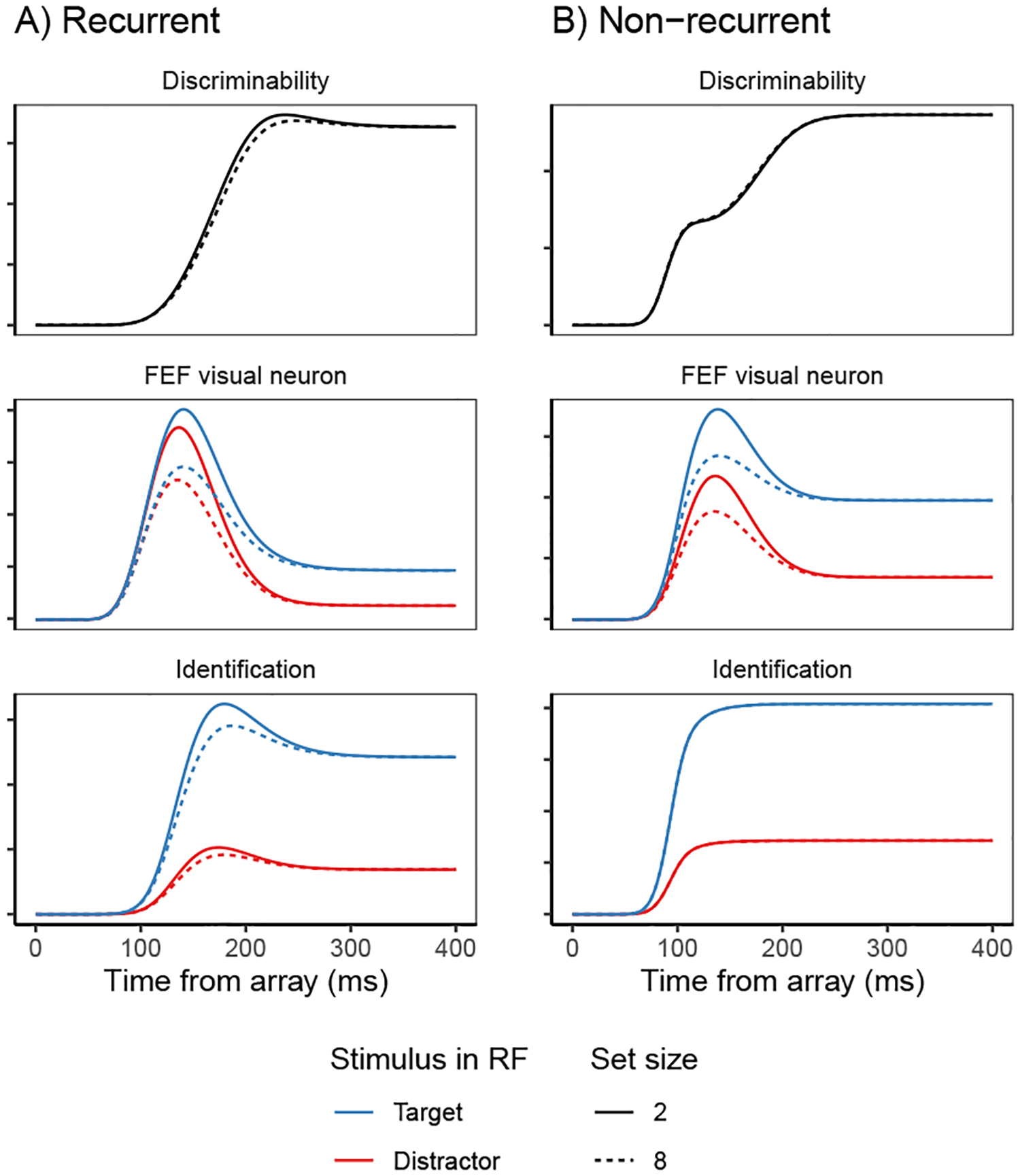
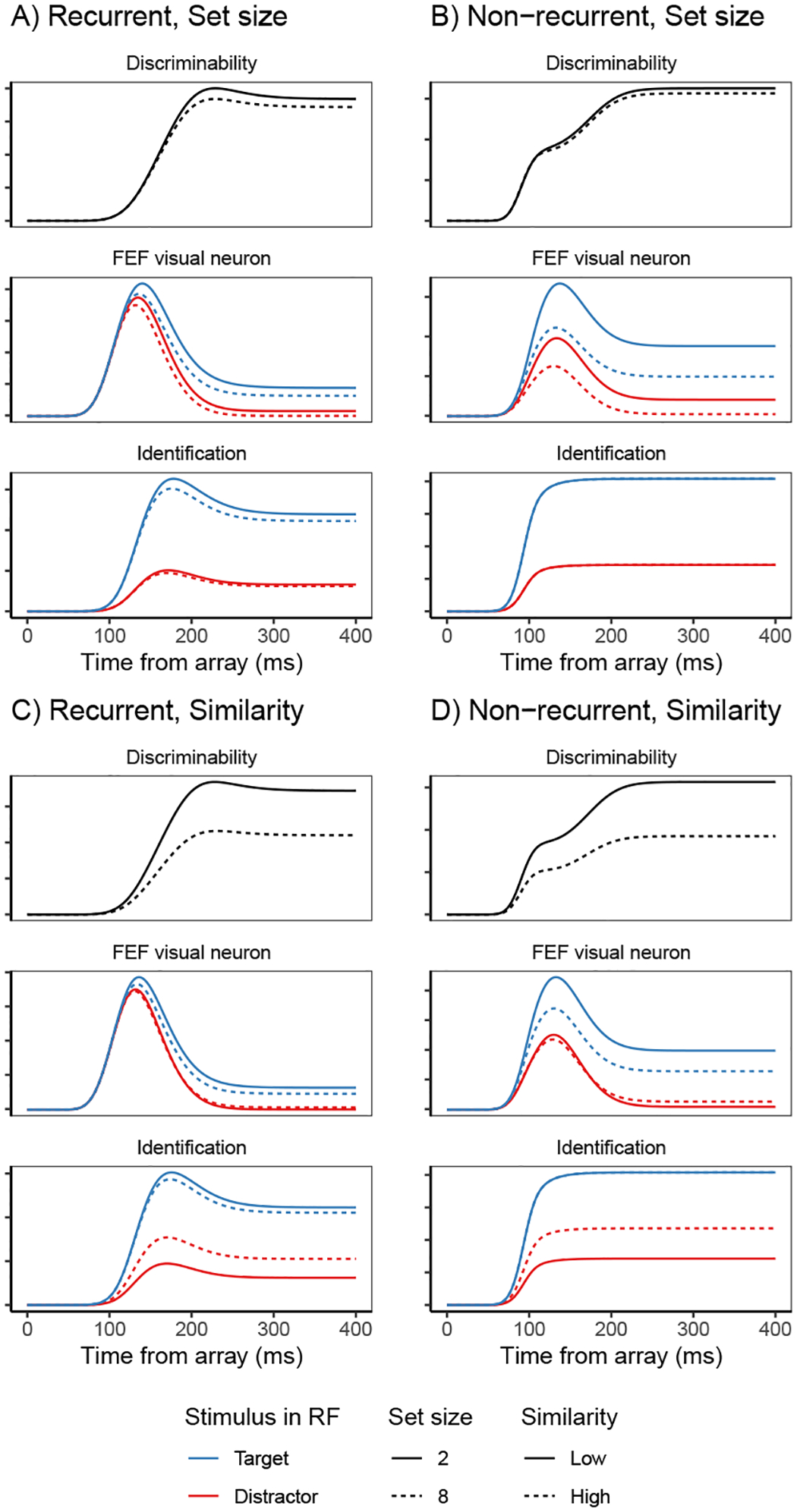
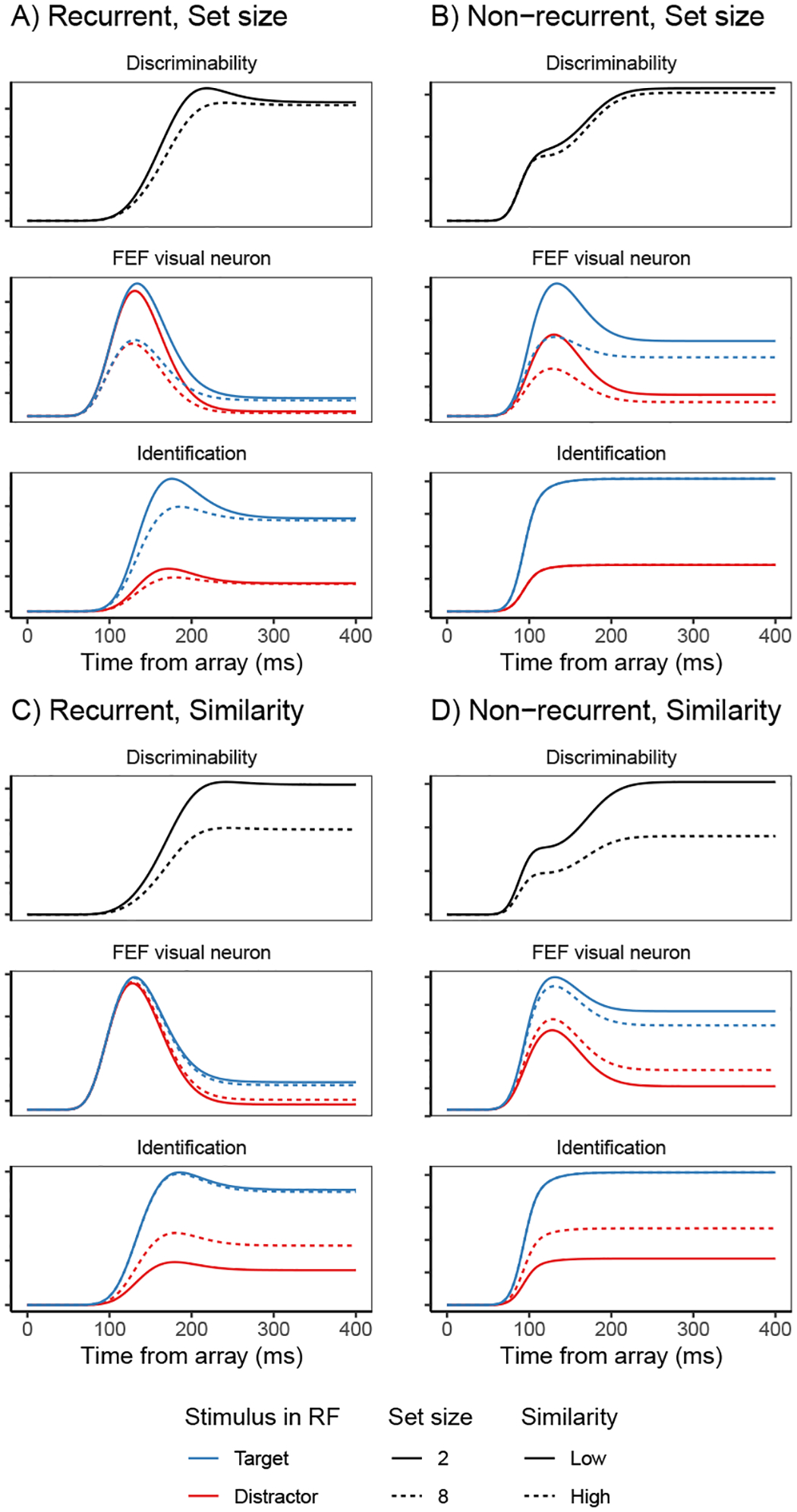
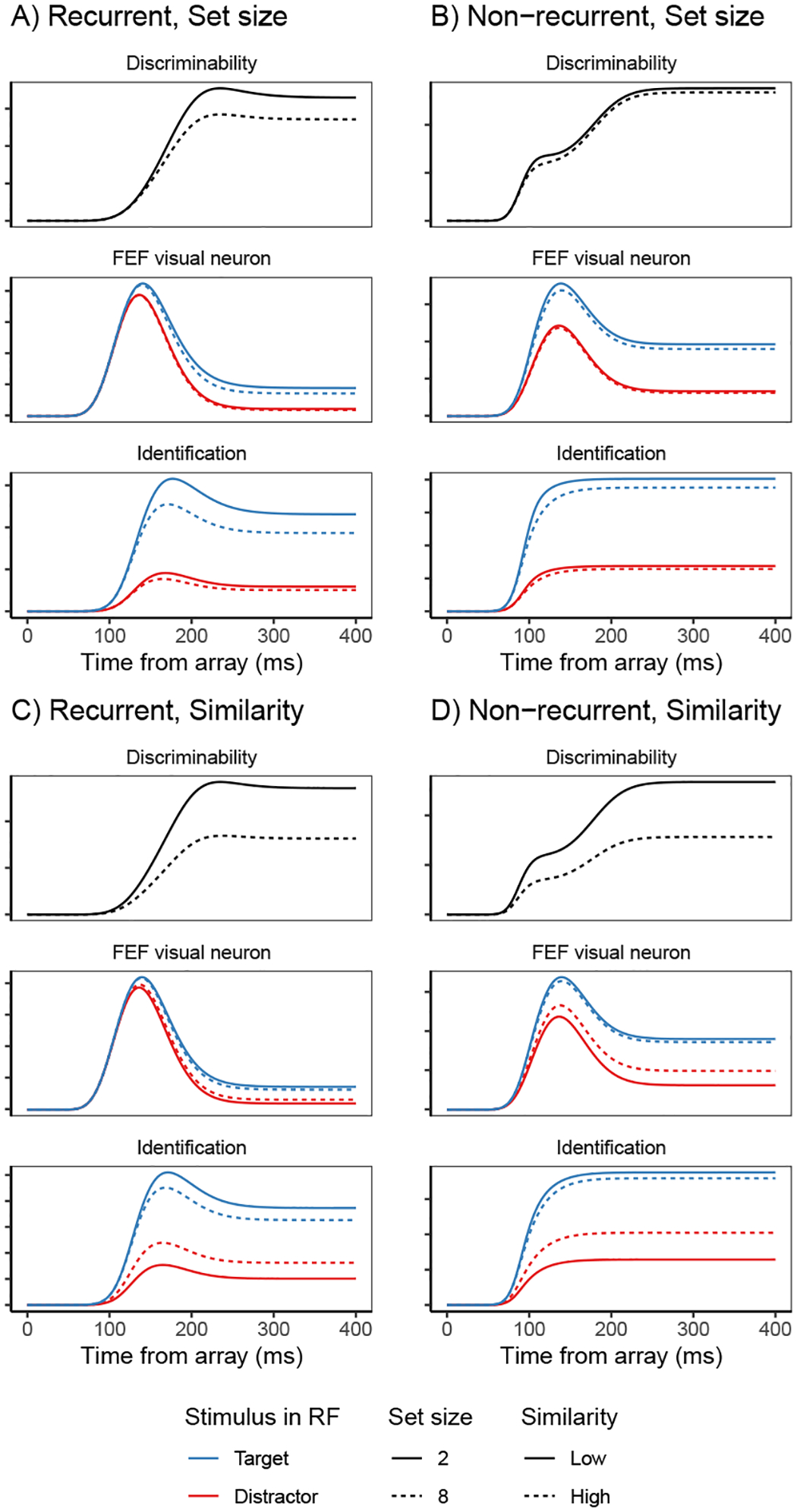
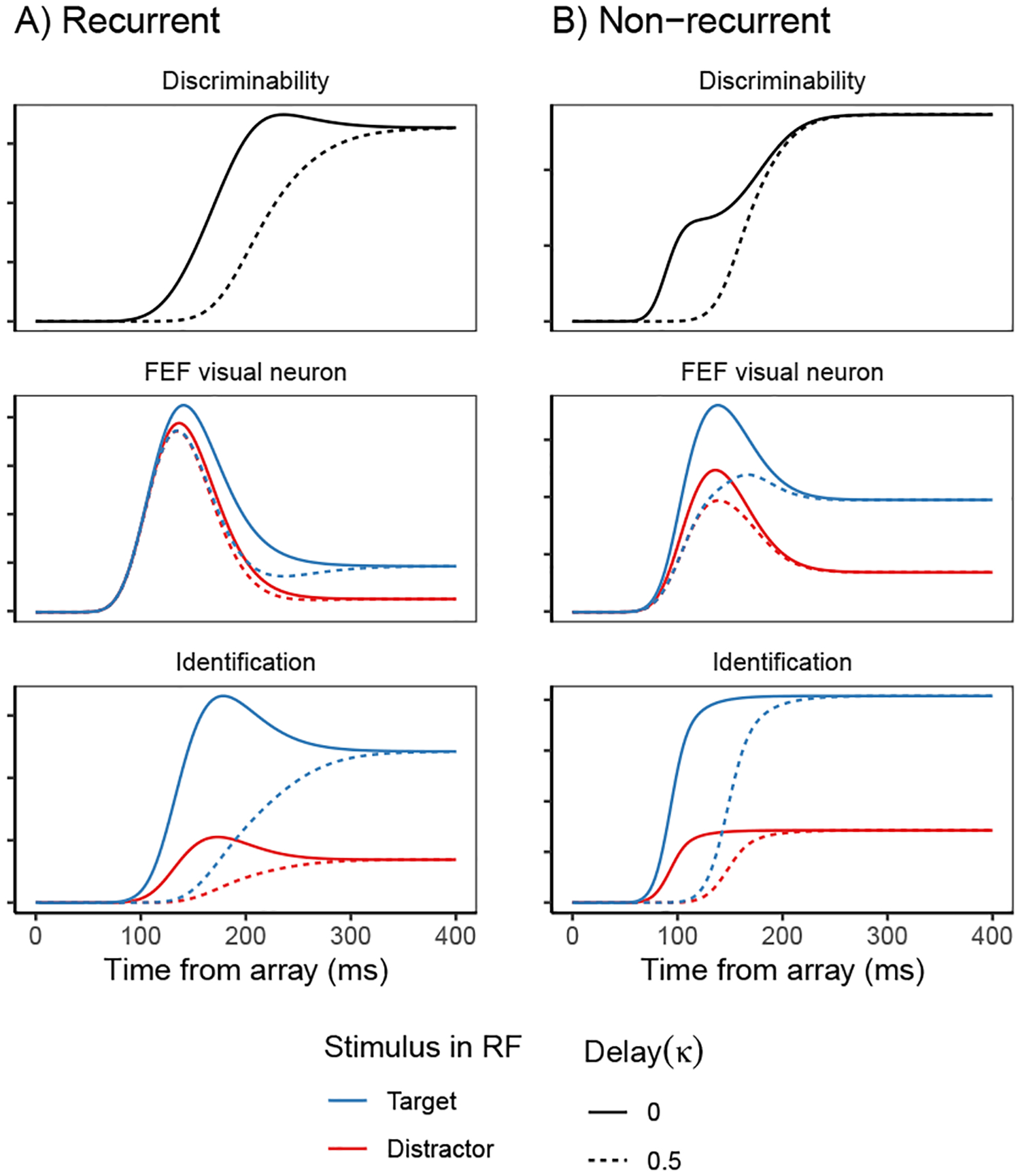
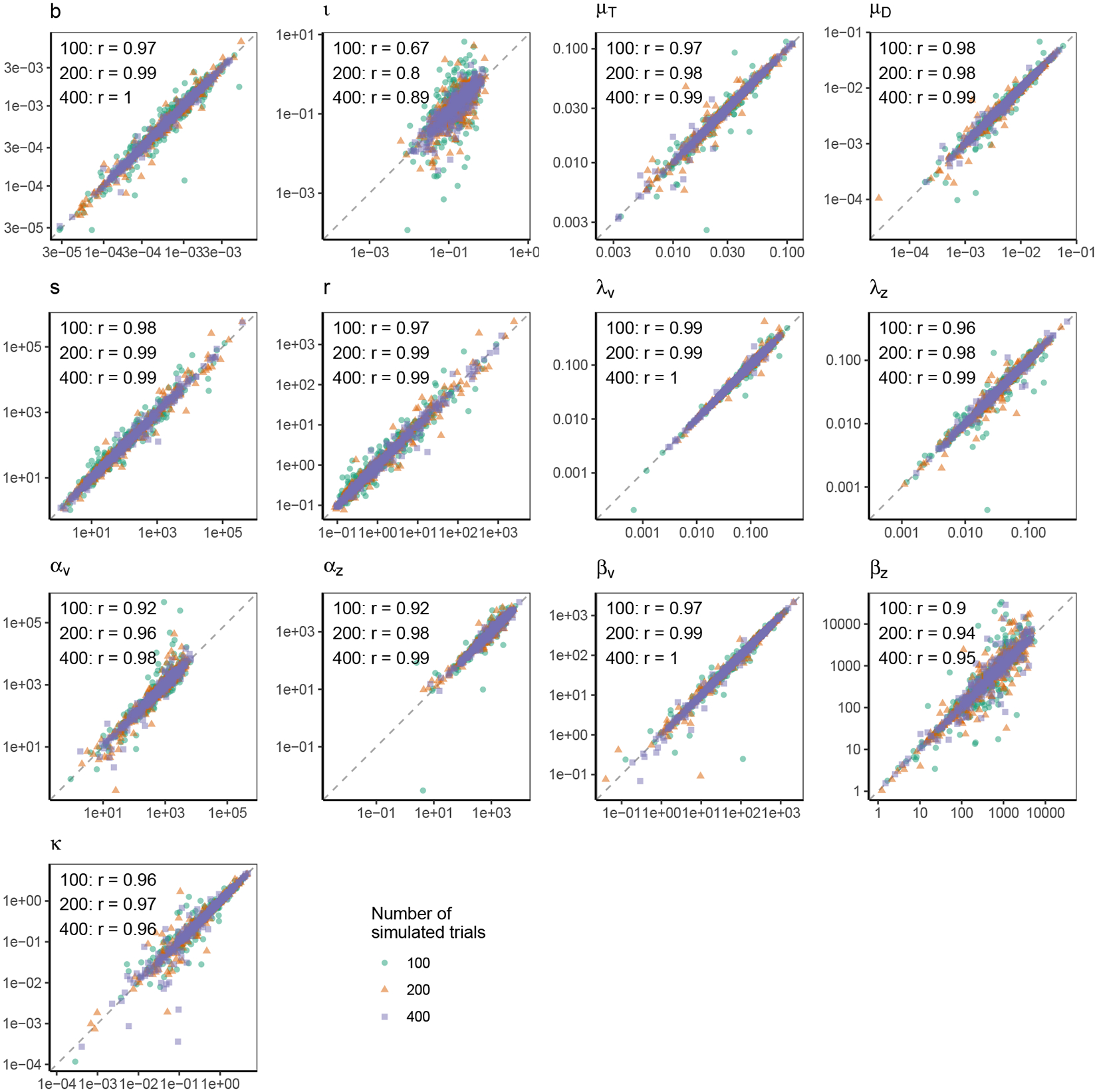
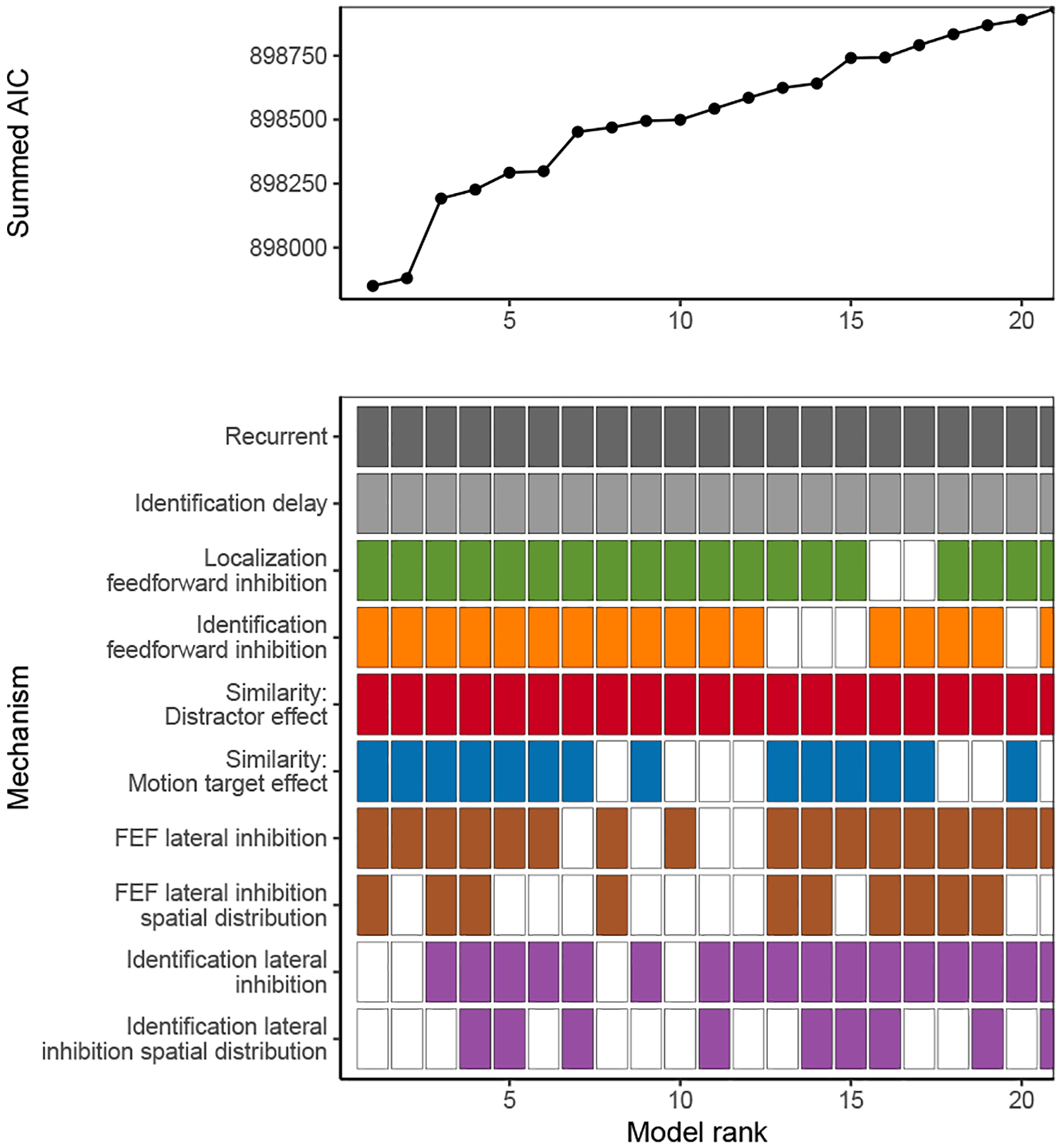
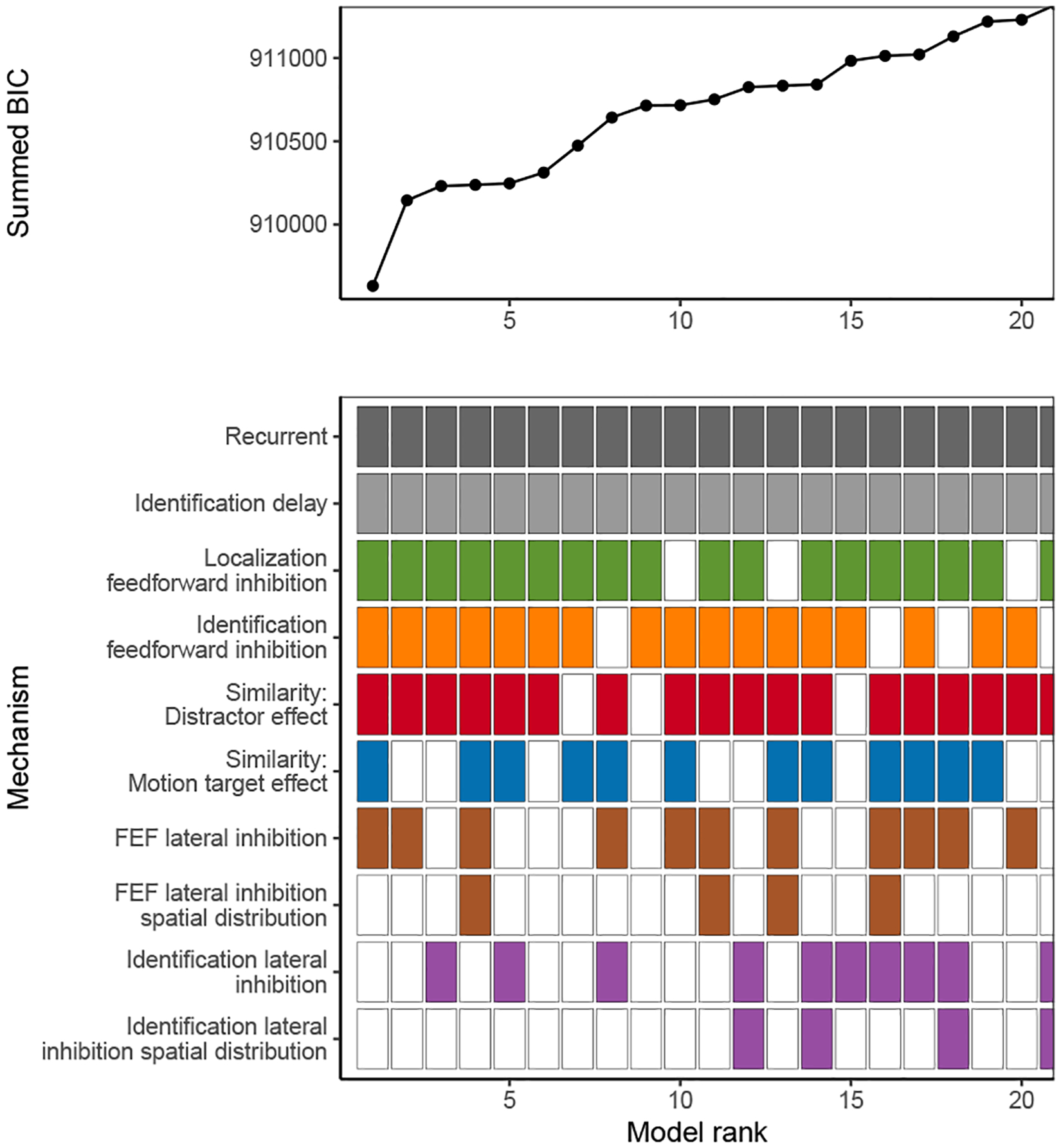
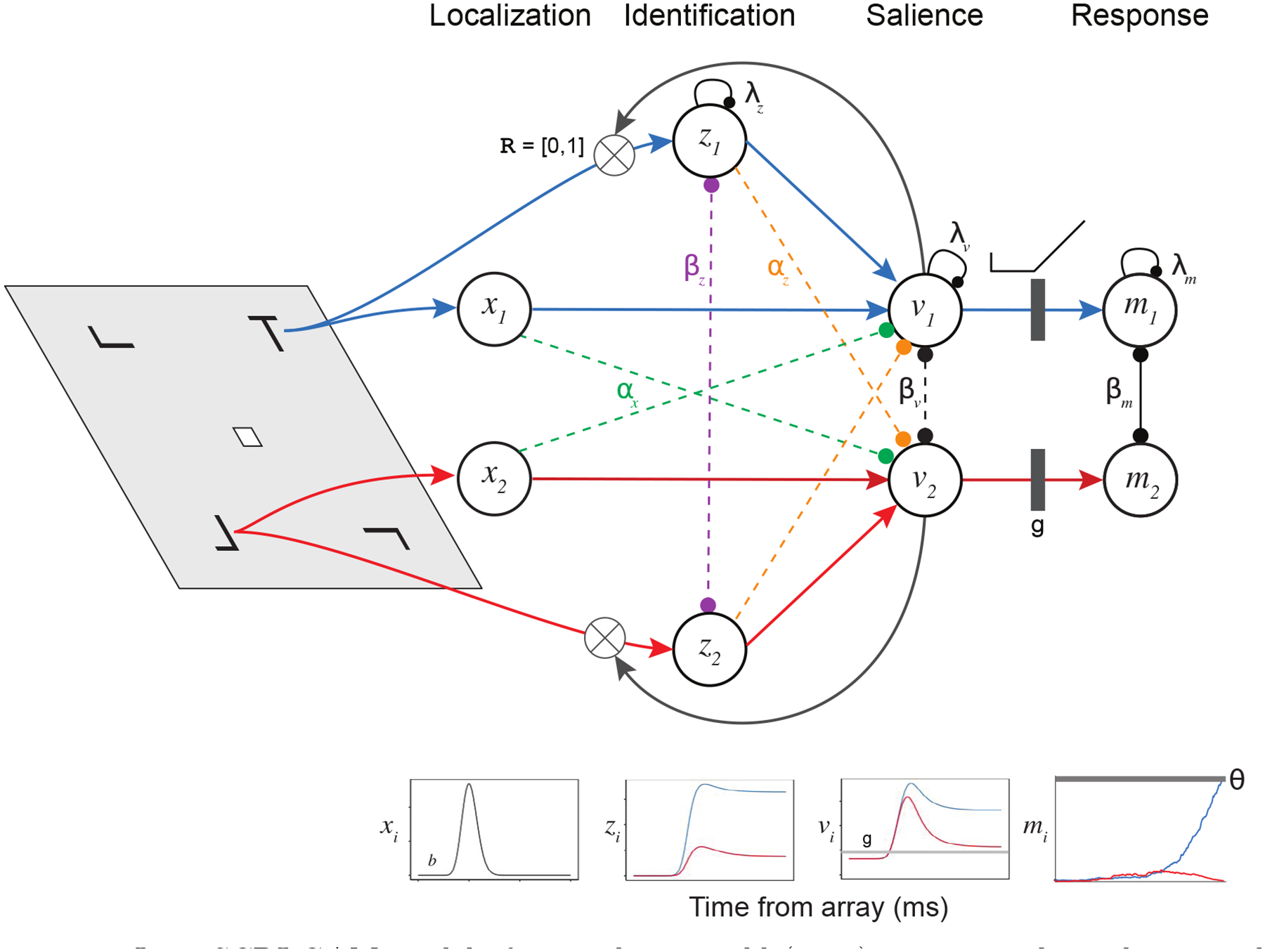
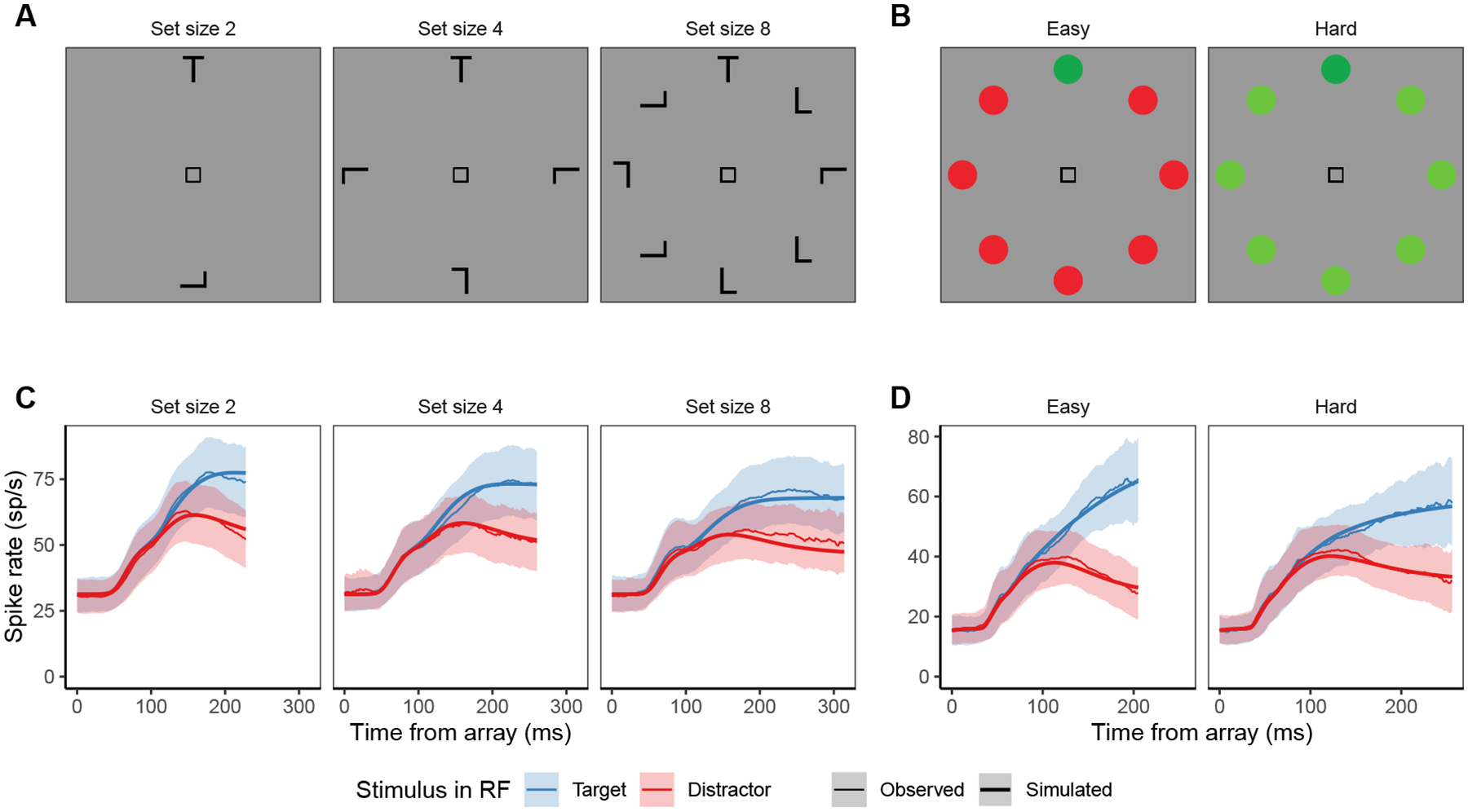
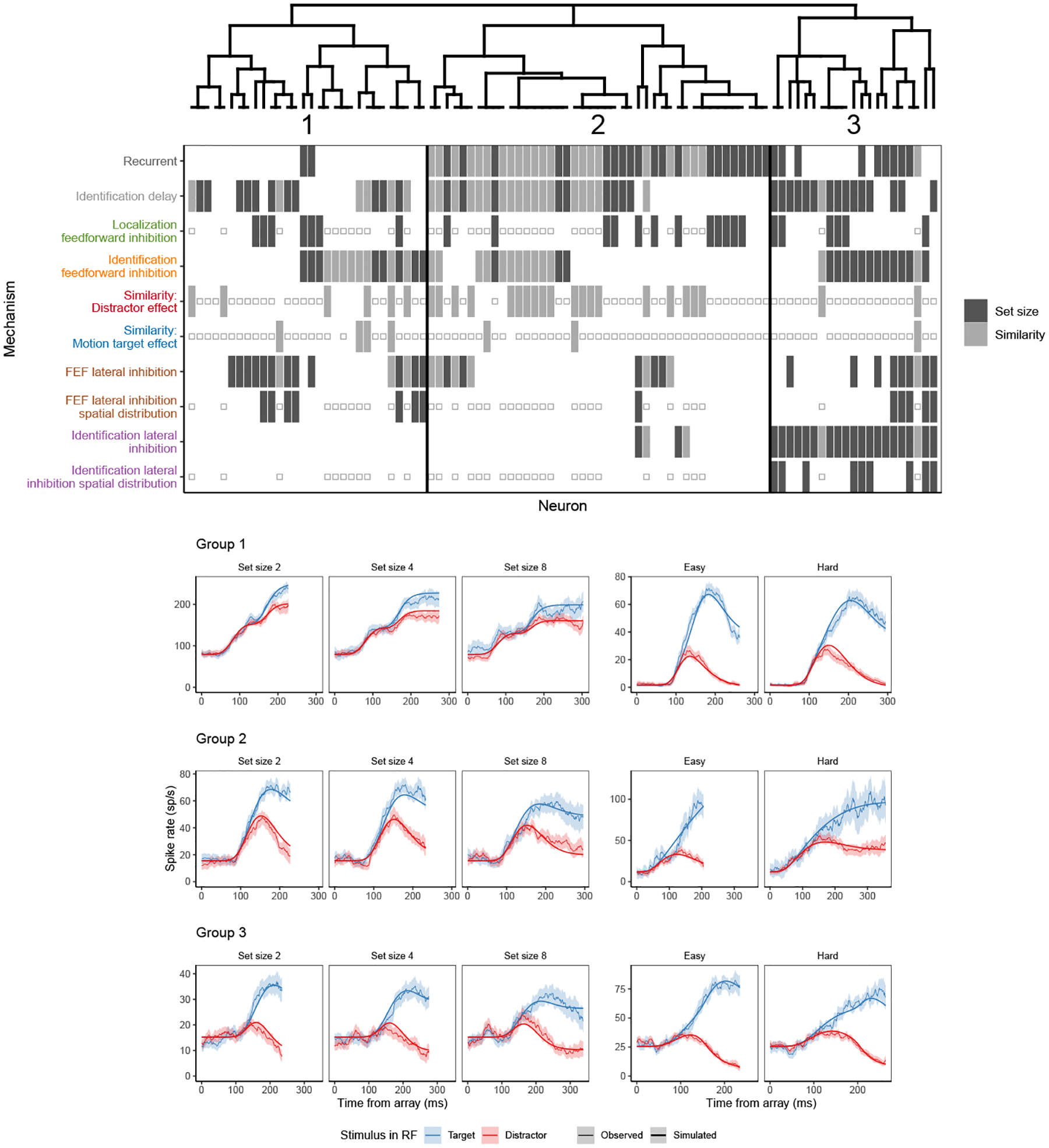
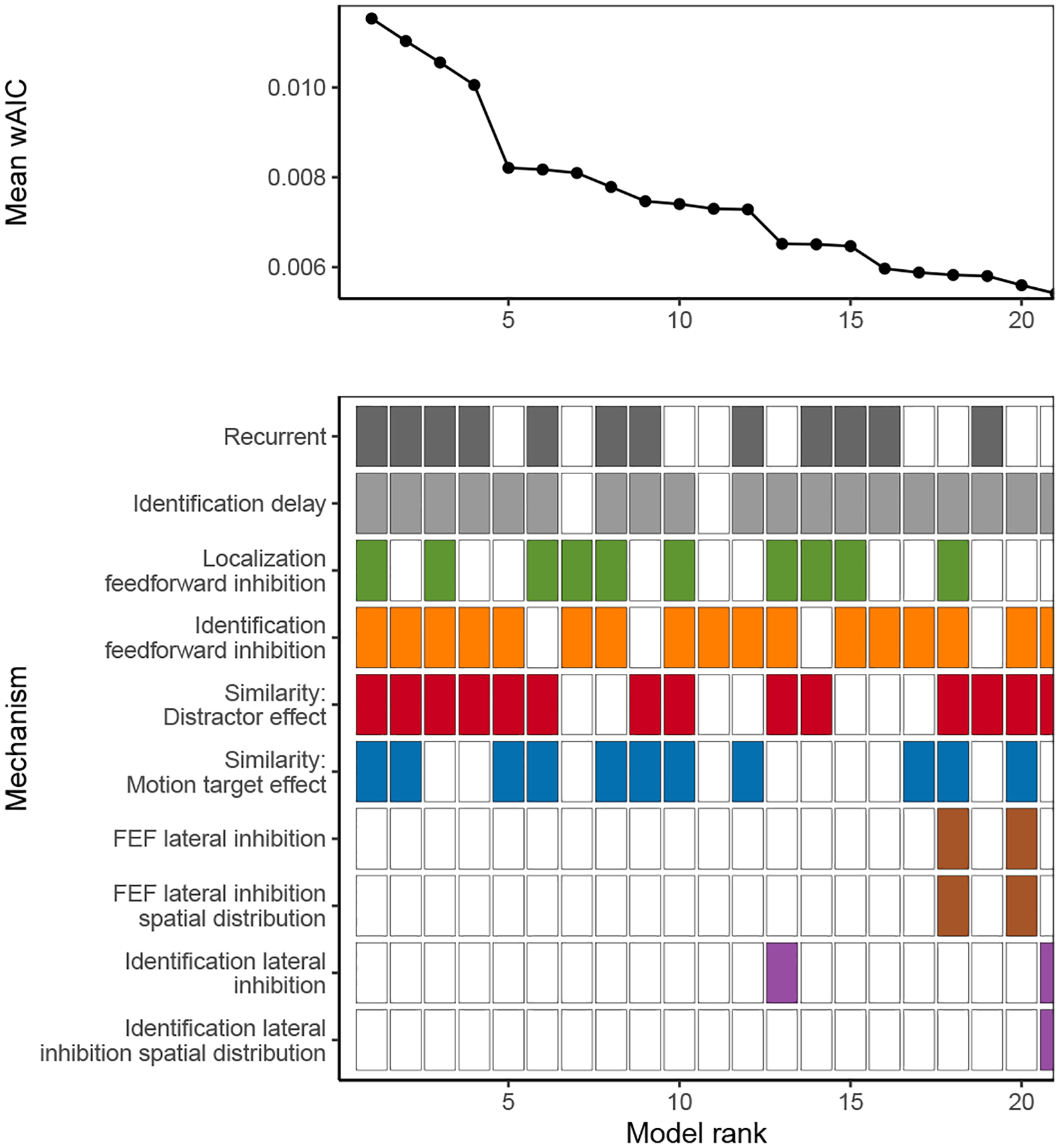
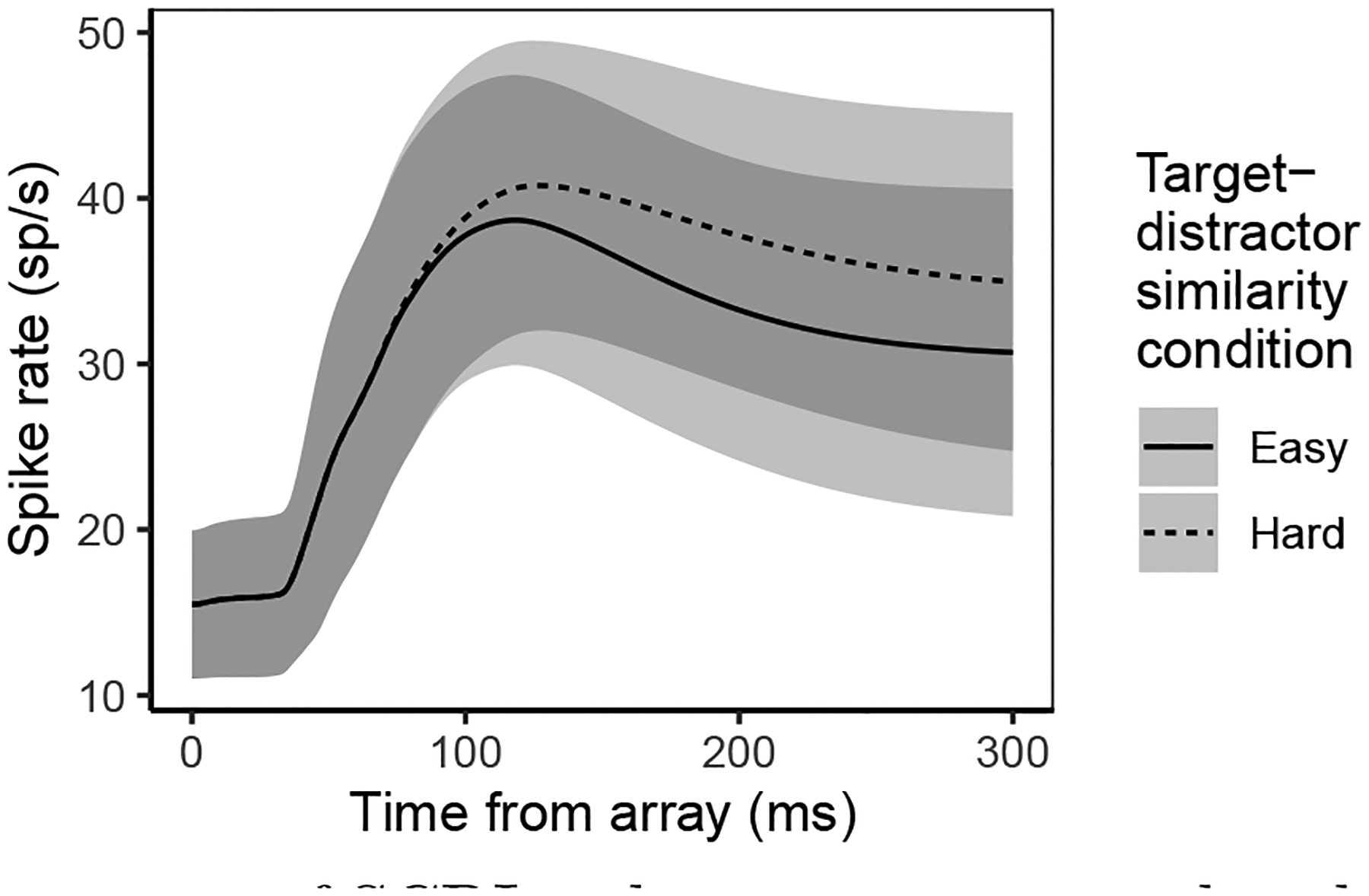
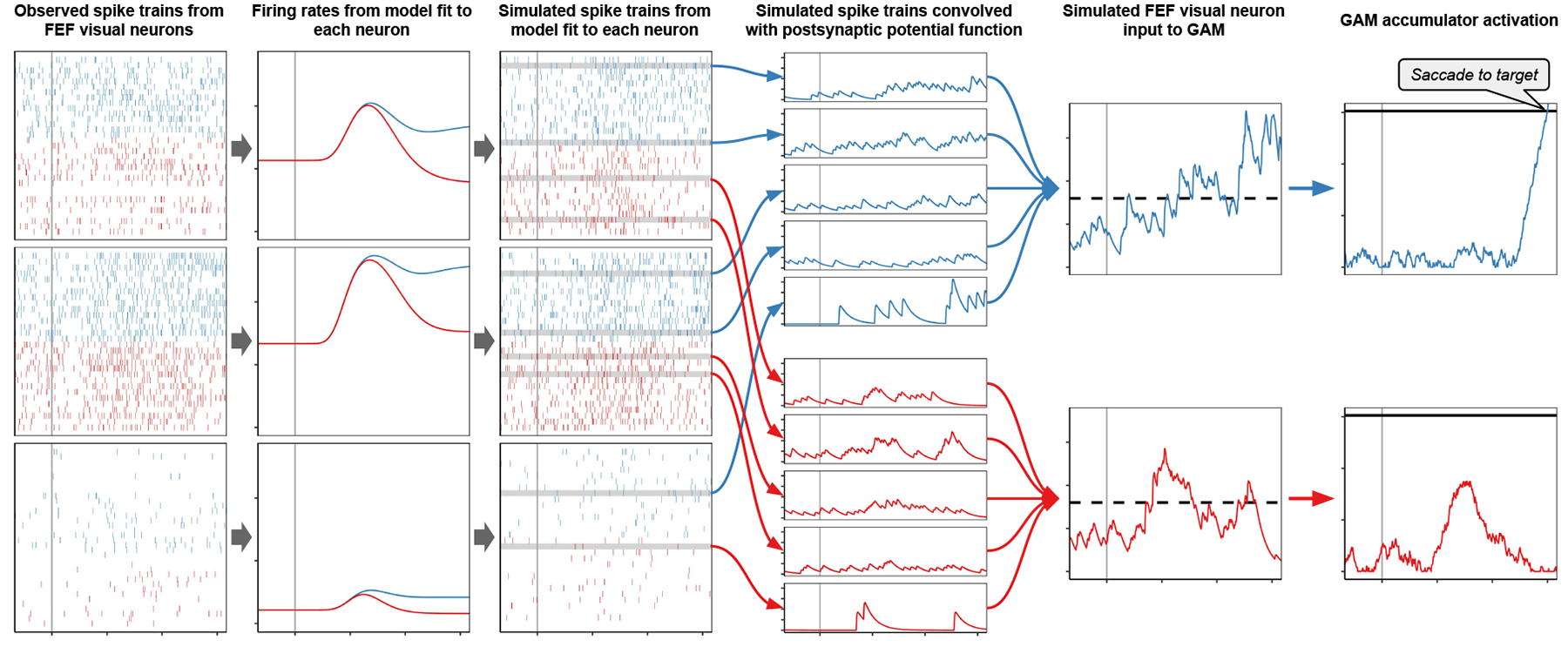
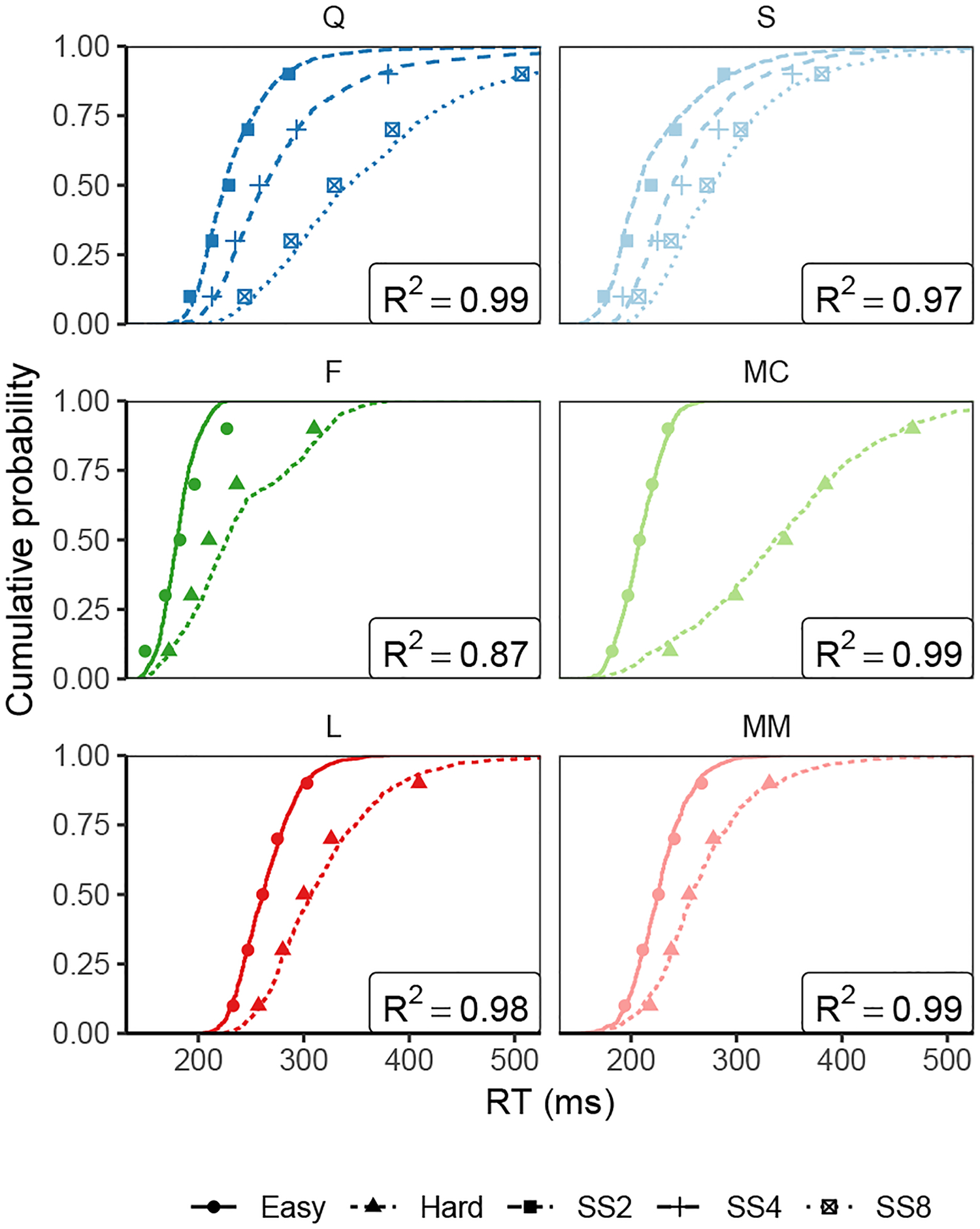
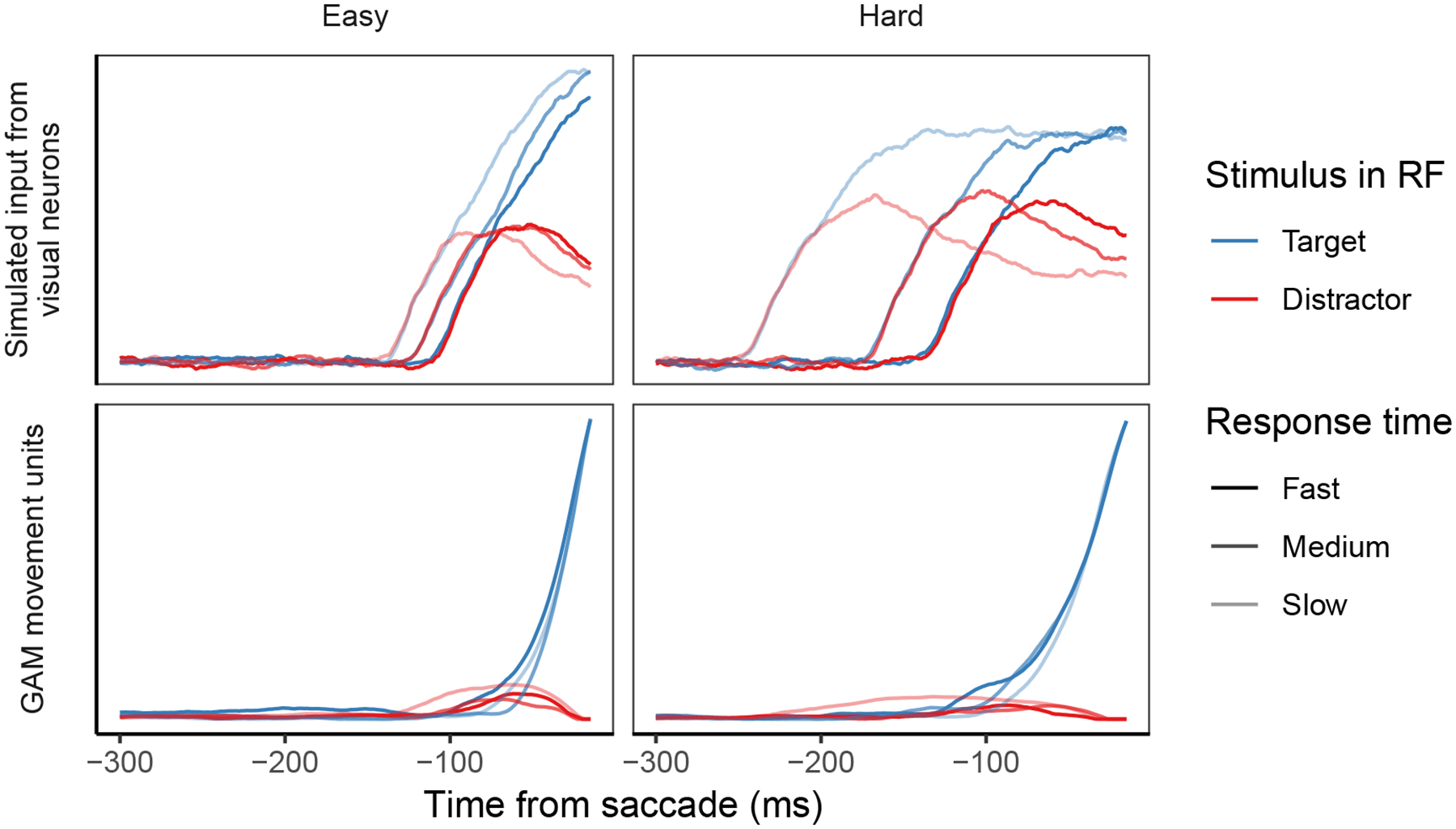
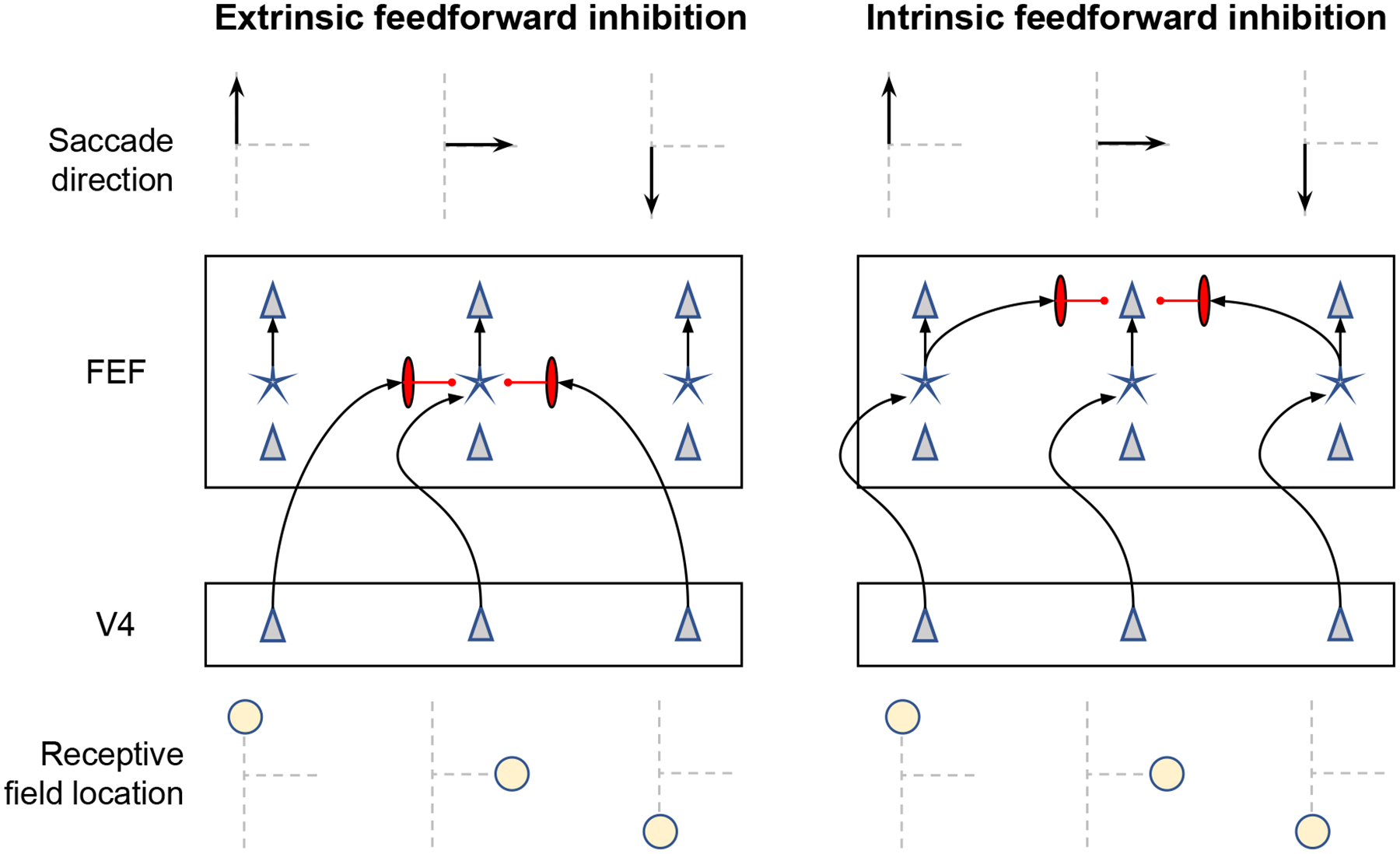
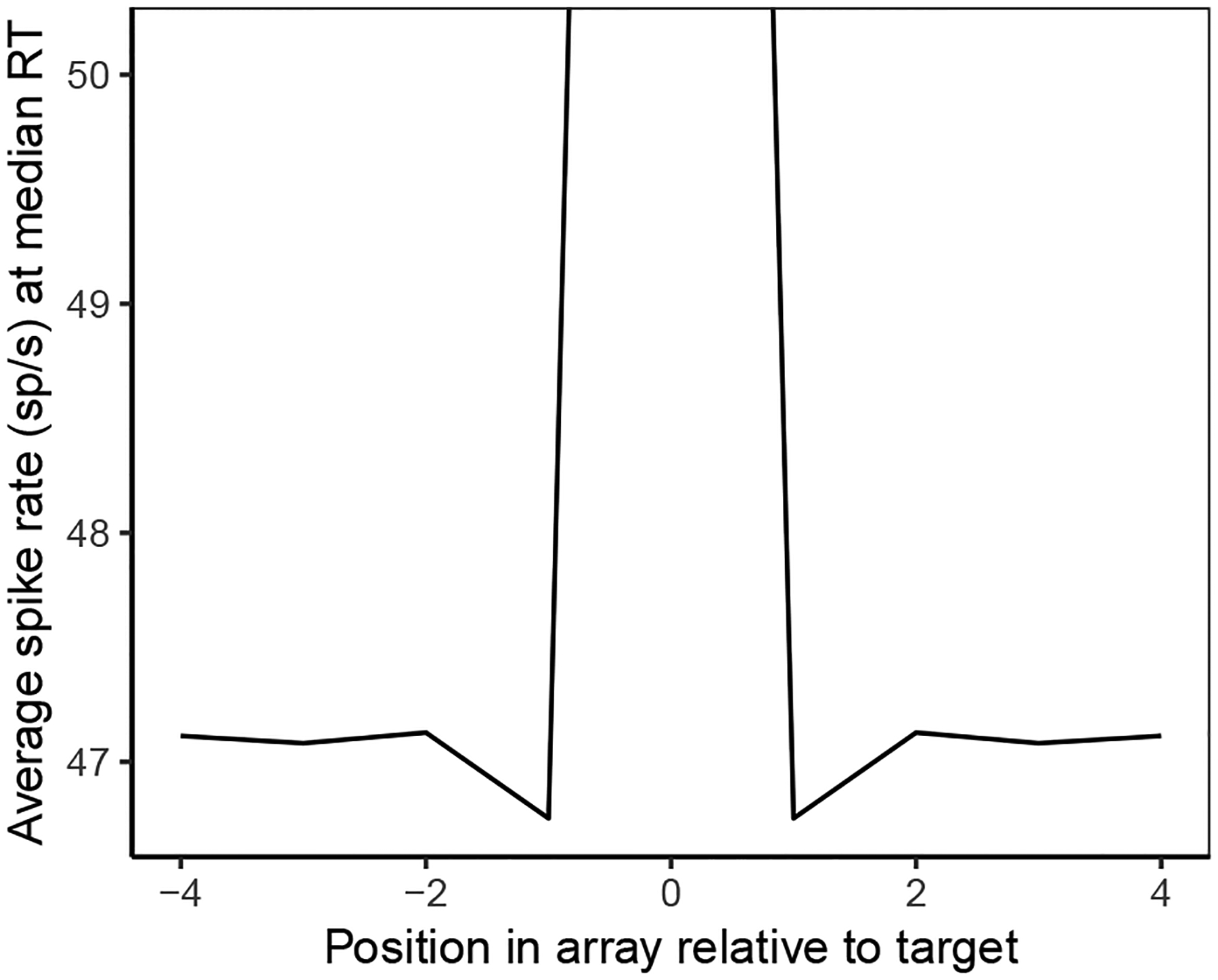
Decisions about where to move the eyes depend on neurons in frontal eye field (FEF). Movement neurons in FEF accumulate salience evidence derived from FEF visual neurons to select the location of a saccade target among distractors. How visual neurons achieve this salience representation is unknown. We present a neuro-computational model of target selection called salience by competitive and recurrent interactions (SCRI), based on the competitive interaction model of attentional selection and decision-making (Smith & Sewell, 2013). SCRI selects targets by synthesizing localization and identification information to yield a dynamically evolving representation of salience across the visual field. SCRI accounts for neural spiking of individual FEF visual neurons, explaining idiosyncratic differences in neural dynamics with specific parameters. Many visual neurons resolve the competition between search items through feedforward inhibition between signals representing different search items, some also require lateral inhibition, and many act as recurrent gates to modulate the incoming flow of information about stimulus identity. SCRI was tested further by using simulated spiking representations of visual salience as input to the gated accumulator model of FEF movement neurons (Purcell et al., 2010, 2012). Predicted saccade response times fit those observed for search arrays of different set sizes and different target-distractor similarities, and accumulator trajectories replicated movement neuron discharge rates. These findings offer new insights into visual decision-making through converging neuro-computational constraints and provide a novel computational account of the diversity of FEF visual neurons. (PsycInfo Database Record (c) 2022 APA, all rights reserved).
Figures
Similar articles
-
Frontal eye field activity before visual search errors reveals the integration of bottom-up and top-down salience.J Neurophysiol. 2005 Jan;93(1):337-51. doi: 10.1152/jn.00330.2004. Epub 2004 Aug 18. J Neurophysiol. 2005. PMID: 15317836 Free PMC article.
-
Perceptual and motor processing stages identified in the activity of macaque frontal eye field neurons during visual search.J Neurophysiol. 1996 Dec;76(6):4040-55. doi: 10.1152/jn.1996.76.6.4040. J Neurophysiol. 1996. PMID: 8985899
-
Response variability of frontal eye field neurons modulates with sensory input and saccade preparation but not visual search salience.J Neurophysiol. 2012 Nov;108(10):2737-50. doi: 10.1152/jn.00613.2012. Epub 2012 Sep 5. J Neurophysiol. 2012. PMID: 22956785 Free PMC article.
-
Neural mechanisms of saccade target selection: gated accumulator model of the visual-motor cascade.Eur J Neurosci. 2011 Jun;33(11):1991-2002. doi: 10.1111/j.1460-9568.2011.07715.x. Eur J Neurosci. 2011. PMID: 21645095 Free PMC article. Review.
-
Neural basis of saccade target selection.Rev Neurosci. 1995 Jan-Mar;6(1):63-85. doi: 10.1515/revneuro.1995.6.1.63. Rev Neurosci. 1995. PMID: 7633641 Review.
Cited by
-
Relating a Spiking Neural Network Model and the Diffusion Model of Decision-Making.Comput Brain Behav. 2022 Sep;5(3):279-301. doi: 10.1007/s42113-022-00143-4. Epub 2022 Jun 13. Comput Brain Behav. 2022. PMID: 36408474 Free PMC article.
-
Vision of objects happens faster and earlier for location than for identity.iScience. 2024 Dec 27;28(2):111702. doi: 10.1016/j.isci.2024.111702. eCollection 2025 Feb 21. iScience. 2024. PMID: 39898052 Free PMC article.
References
-
- Akaike H (1974). A new look at the statistical model identification. IEEE Transactions on Automatic Control, 19(6), 716–723. doi: 10.1109/TAC.1974.1100705 - DOI
-
- Atkinson RC, Holmgren JE, & Juola JF (1969). Processing time as influenced by the number of elements in a visual display. Perception & Psychophysics, 6(6), 321–326. doi: 10.3758/BF03212784 - DOI
