

Microbial Protein Binding to gC1qR Drives PLA2G1B-Induced CD4 T-Cell Anergy
- PMID: 35392090
- PMCID: PMC8981723
- DOI: 10.3389/fimmu.2022.824746
Microbial Protein Binding to gC1qR Drives PLA2G1B-Induced CD4 T-Cell Anergy
Abstract
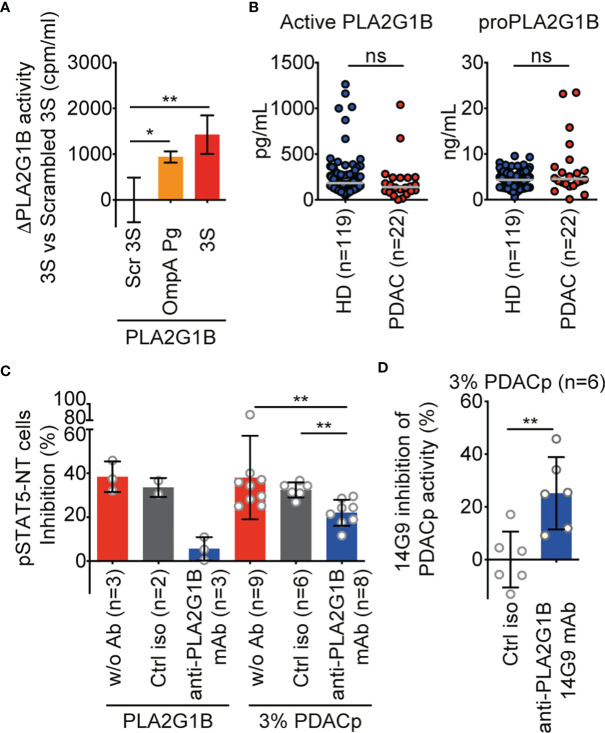
The origin of the impaired CD4 T-cell response and immunodeficiency of HIV-infected patients is still only partially understood. We recently demonstrated that PLA2G1B phospholipase synergizes with the HIV gp41 envelope protein in HIV viremic plasma to induce large abnormal membrane microdomains (aMMDs) that trap and inactivate physiological receptors, such as those for IL-7. However, the mechanism of regulation of PLA2G1B activity by the cofactor gp41 is not known. Here, we developed an assay to directly follow PLA2G1B enzymatic activity on CD4 T-cell membranes. We demonstrated that gp41 directly binds to PLA2G1B and increases PLA2G1B enzymatic activity on CD4 membrane. Furthermore, we show that the conserved 3S sequence of gp41, known to bind to the innate sensor gC1qR, increases PLA2G1B activity in a gC1qR-dependent manner using gC1qR KO cells. The critical role of the 3S motif and gC1qR in the inhibition of CD4 T-cell function by the PLA2G1B/cofactor system in HIV-infected patients led us to screen additional microbial proteins for 3S-like motifs and to study other proteins known to bind to the gC1qR to further investigate the role of the PLA2G1B/cofactor system in other infectious diseases and carcinogenesis. We have thus extended the PLA2G1B/cofactor system to HCV and Staphylococcus aureus infections and additional pathologies where microbial proteins with 3S-like motifs also increase PLA2G1B enzymatic activity. Notably, the bacteria Porphyromonas gingivalis, which is associated with pancreatic ductal adenocarcinoma (PDAC), encodes such a cofactor protein and increased PLA2G1B activity in PDAC patient plasma inhibits the CD4 response to IL-7. Our findings identify PLA2G1B/cofactor system as a CD4 T-cell inhibitor. It involves the gC1qR and disease-specific cofactors which are gC1qR-binding proteins that can contain 3S-like motifs. This mechanism involved in HIV-1 immunodeficiency could play a role in pancreatic cancer and several other diseases. These observations suggest that the PLA2G1B/cofactor system is a general CD4 T-cell inhibitor and pave the way for further studies to better understand the role of CD4 T-cell anergy in infectious diseases and tumor escape.
Keywords: CD4 T cell; HCV; HIV; PLA2G1B; gC1qR; infectious disease; porphyromonas gingivalis; staphylococcus aureus.
Copyright © 2022 Pothlichet, Meola, Bugault, Jeammet, Savitt, Ghebrehiwet, Touqui, Pouletty, Fiore, Sauvanet and Thèze.
Conflict of interest statement
JT is cofounder and CSO of DIACCURATE, a spin-off of the Institut Pasteur. JP, AM, FB, and LJ are, or were, employees of DIACCURATE. PP was employed by company Truffle Capital. BG receives royalties from the sale of anti-gC1qR antibodies and gC1qR detection assay kit. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
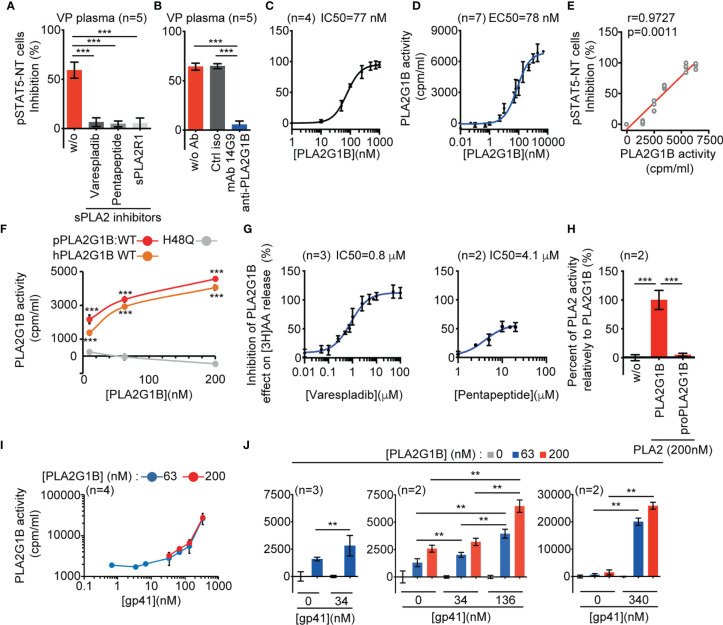
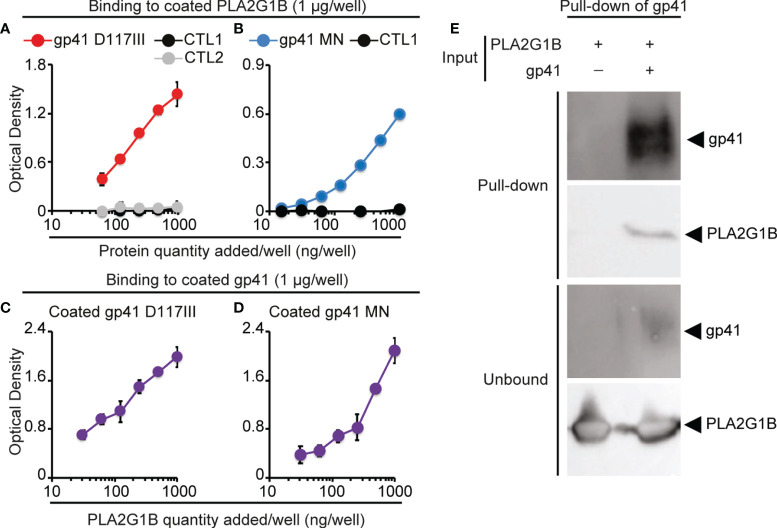
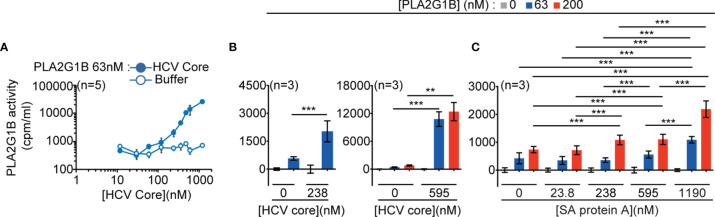
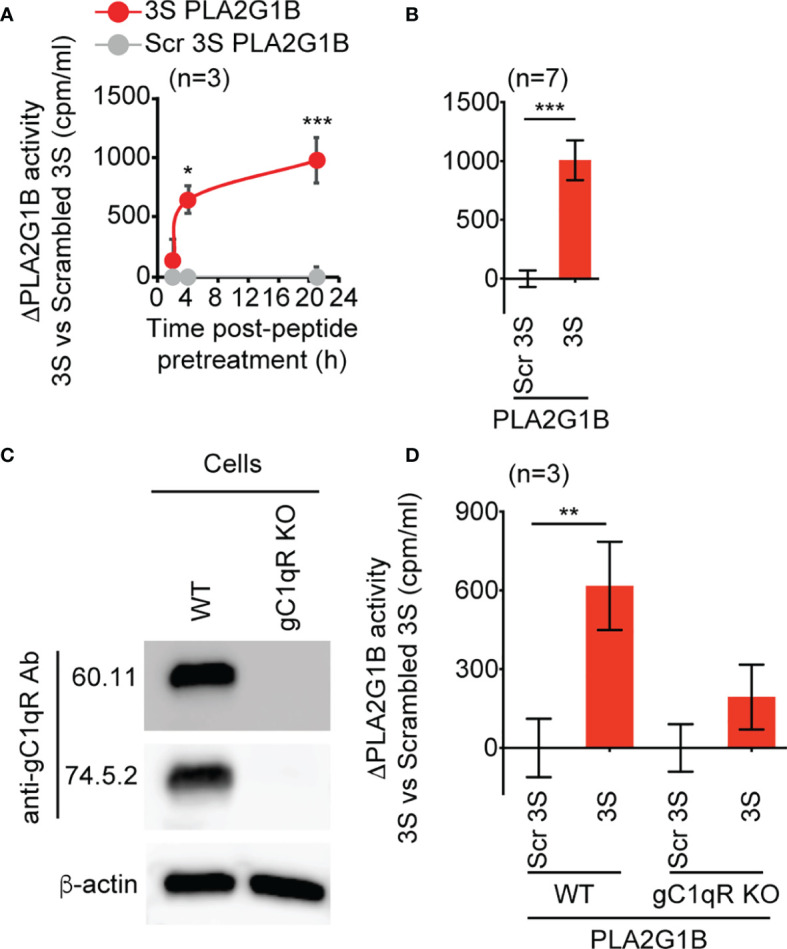
References
-
- Pednekar L, Valentino A, Ji Y, Tumma N, Valentino C, Kadoor A, et al. . Identification of the Gc1qr Sites for the HIV-1 Viral Envelope Protein Gp41 and the HCV Core Protein: Implications in Viral-Specific Pathogenesis and Therapy. Mol Immunol (2016) 74:18–26. doi: 10.1016/J.MOLIMM.2016.03.016 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials