

Catastrophic hydraulic failure and tipping points in plants
- PMID: 35394656
- PMCID: PMC9544843
- DOI: 10.1111/pce.14327
Catastrophic hydraulic failure and tipping points in plants
Abstract
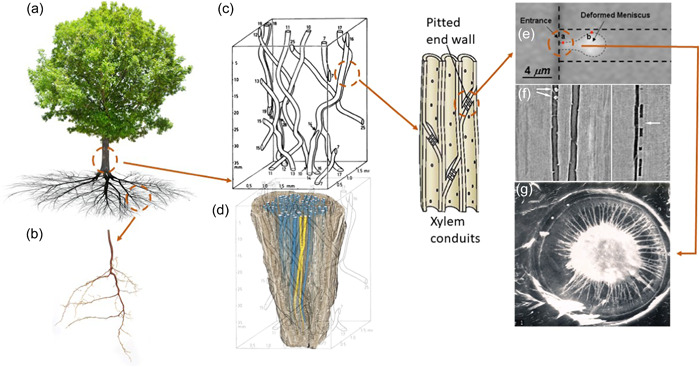
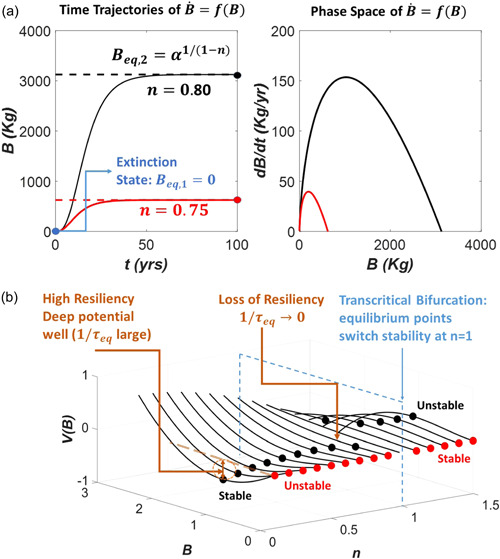
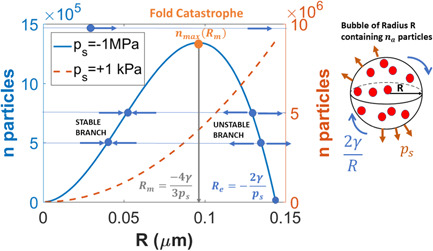
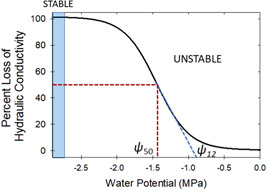
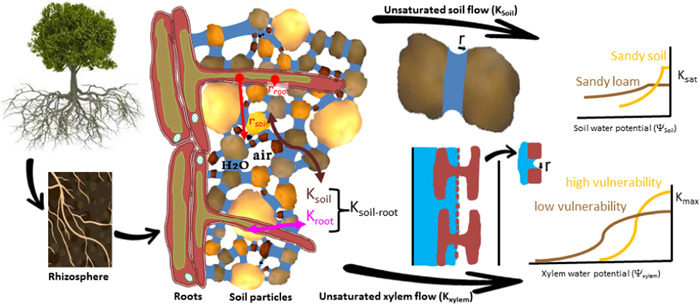
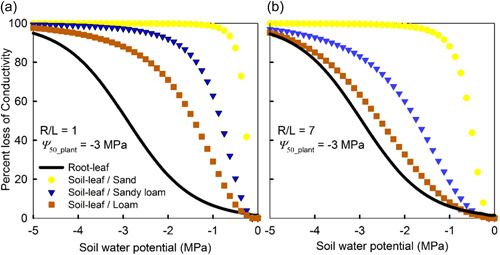
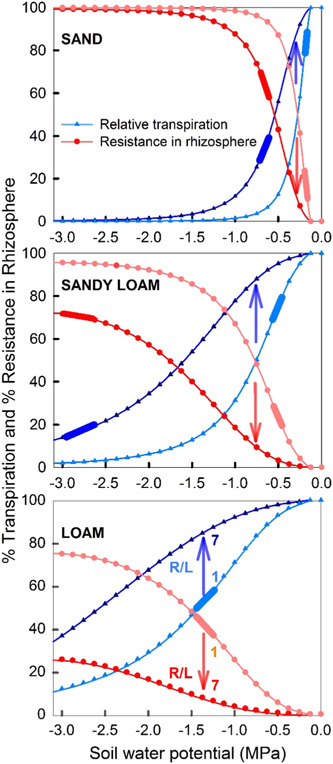
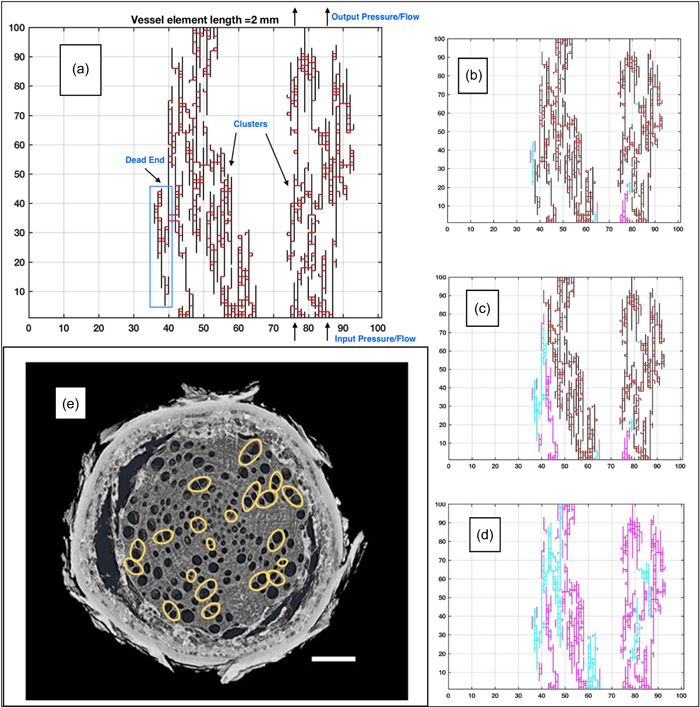
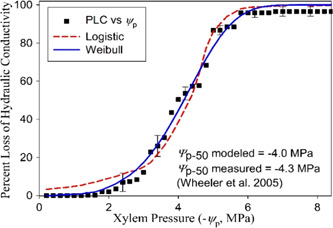
Water inside plants forms a continuous chain from water in soils to the water evaporating from leaf surfaces. Failures in this chain result in reduced transpiration and photosynthesis and are caused by soil drying and/or cavitation-induced xylem embolism. Xylem embolism and plant hydraulic failure share several analogies to 'catastrophe theory' in dynamical systems. These catastrophes are often represented in the physiological and ecological literature as tipping points when control variables exogenous (e.g., soil water potential) or endogenous (e.g., leaf water potential) to the plant are allowed to vary on time scales much longer than time scales associated with cavitation events. Here, plant hydraulics viewed from the perspective of catastrophes at multiple spatial scales is considered with attention to bubble expansion within a xylem conduit, organ-scale vulnerability to embolism, and whole-plant biomass as a proxy for transpiration and hydraulic function. The hydraulic safety-efficiency tradeoff, hydraulic segmentation and maximum plant transpiration are examined using this framework. Underlying mechanisms for hydraulic failure at fine scales such as pit membranes and cell-wall mechanics, intermediate scales such as xylem network properties and at larger scales such as soil-tree hydraulic pathways are discussed. Understudied areas in plant hydraulics are also flagged where progress is urgently needed.
Keywords: bifurcation; cavitation; cusp; embolism; fold; r-shaped curves; s-shaped curves; soil; transpiration; water potential; xylem.
© 2022 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
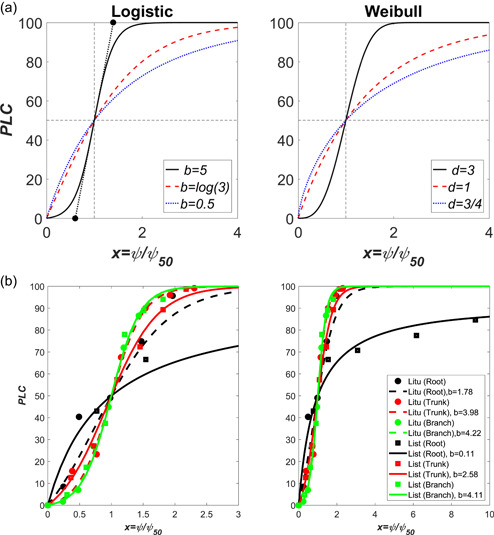
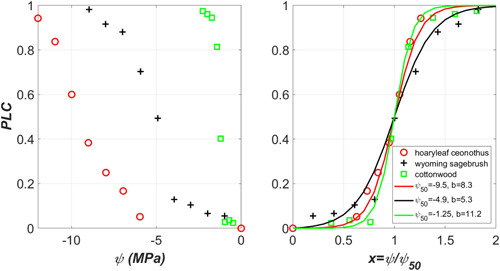
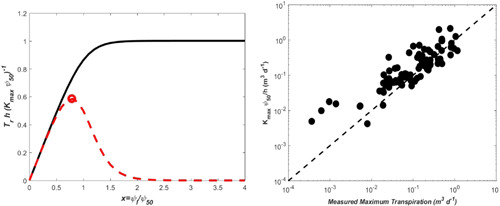
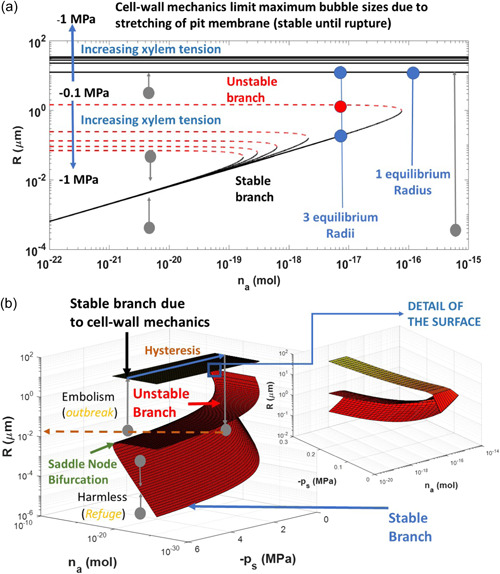
References
-
- Adams, H.D. , Zeppel, M.J. , Anderegg, W.R. , Hartmann, H. , Landhäusser, S.M. , Tissue, D.T. et al. (2017) A multi‐species synthesis of physiological mechanisms in drought‐induced tree mortality. Nature Ecology and Evolution, 1, 1285–1291. - PubMed
-
- Albuquerque, C. , Scoffoni, C. , Brodersen, C.R. , Buckley, T.N. , Sack, L. & McElrone, A.J. (2020) Coordinated decline of leaf hydraulic and stomatal conductances under drought is not linked to leaf xylem embolism for different grapevine cultivars. Journal of Experimental Botany, 71, 7286–7300. - PubMed
-
- Allen, C.D. , Breshears, D.D. & McDowell, N.G. (2015) On underestimation of global vulnerability to tree mortality and forest die‐off from hotter drought in the Anthropocene. Ecosphere, 6, 1–55.
-
- Allen, C.D. , Macalady, A.K. , Chenchouni, H. , Bachelet, D. , McDowell, N. , Vennetier, M. et al. (2010) A global overview of drought and heat‐induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 259, 660–684.
-
- Anfodillo, T. , Carraro, V. , Carrer, M. , Fior, C. & Rossi, S. (2006) Convergent tapering of xylem conduits in different woody species. New Phytologist, 169, 279–290. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
