

Autoreactive memory Th17 cells are principally derived from T-bet+RORγt+ Th17/1 effectors
- PMID: 35395541
- PMCID: PMC9106930
- DOI: 10.1016/j.jaut.2022.102816
Autoreactive memory Th17 cells are principally derived from T-bet+RORγt+ Th17/1 effectors
Abstract
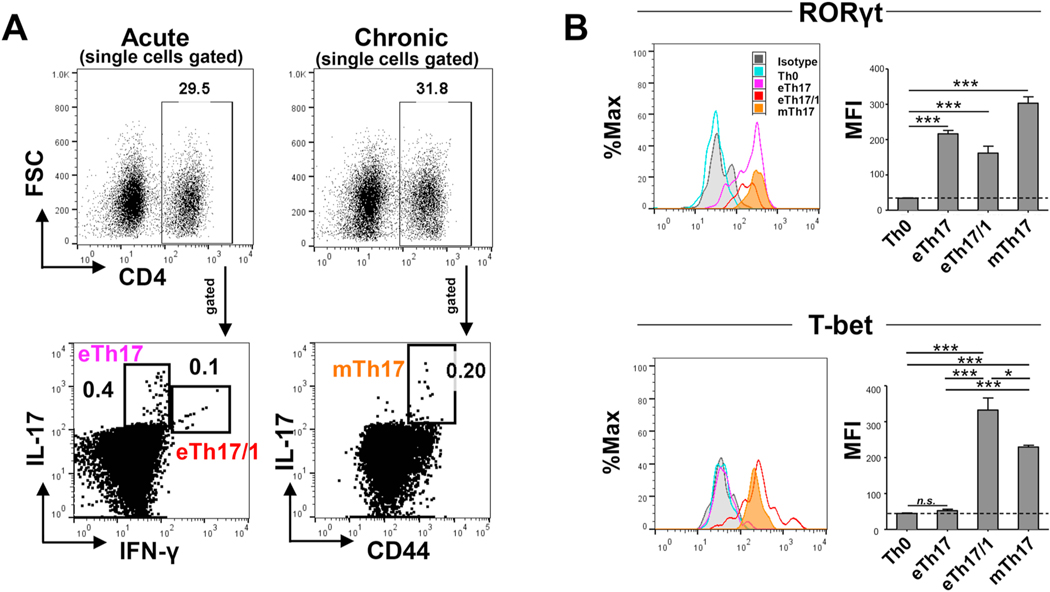
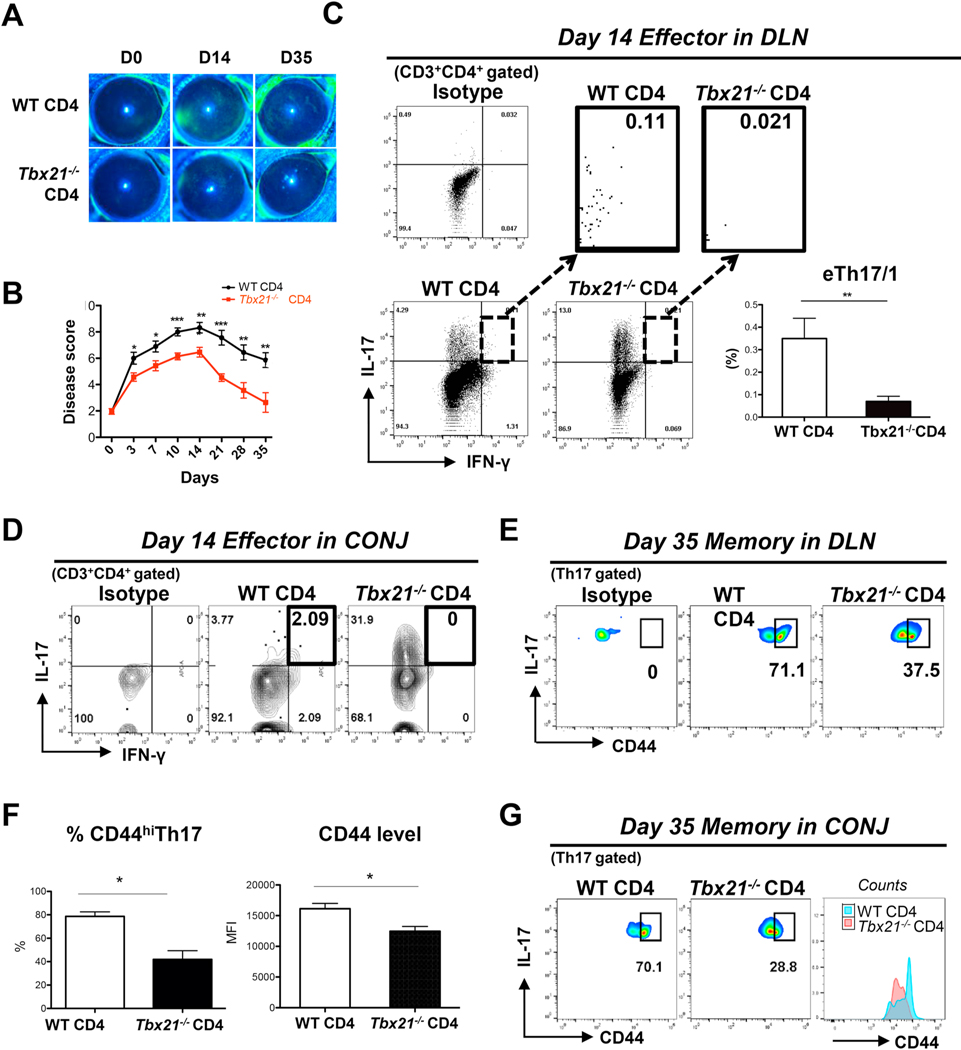
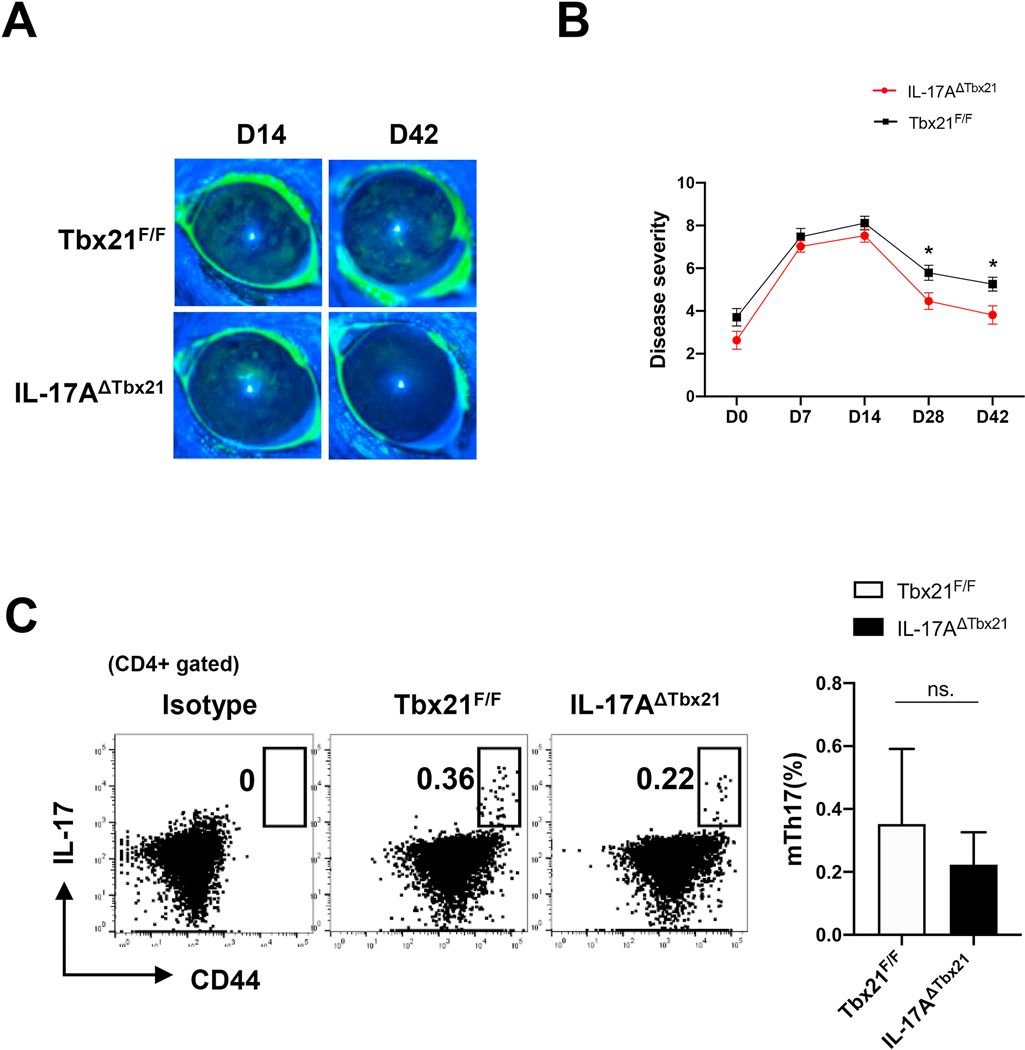
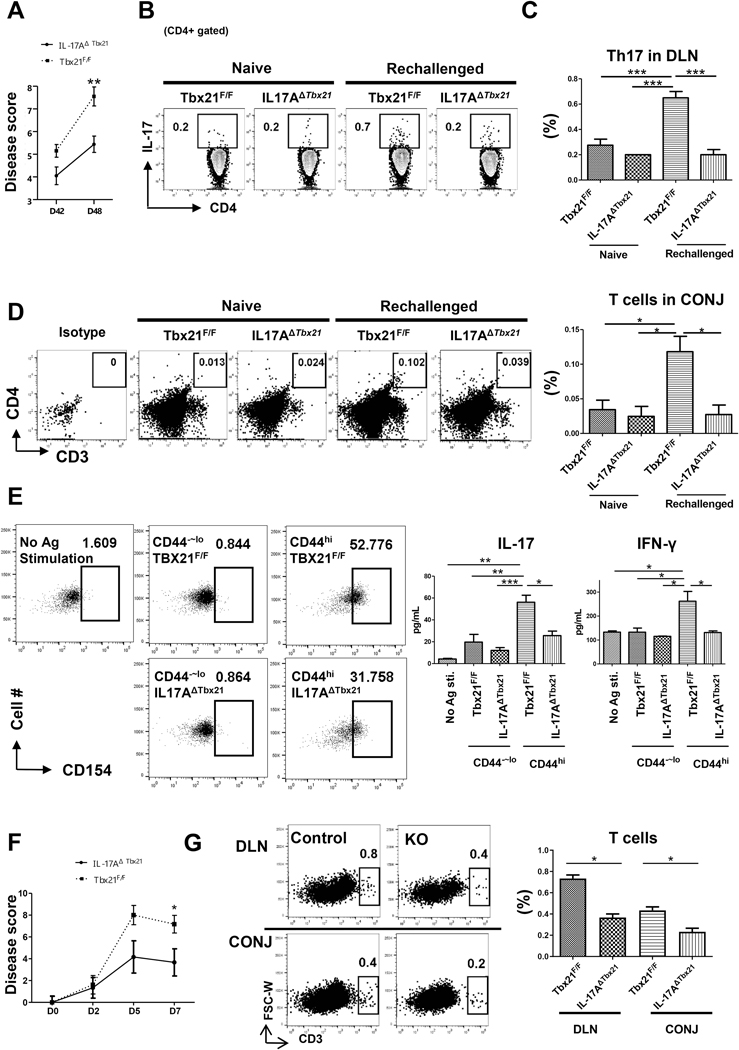
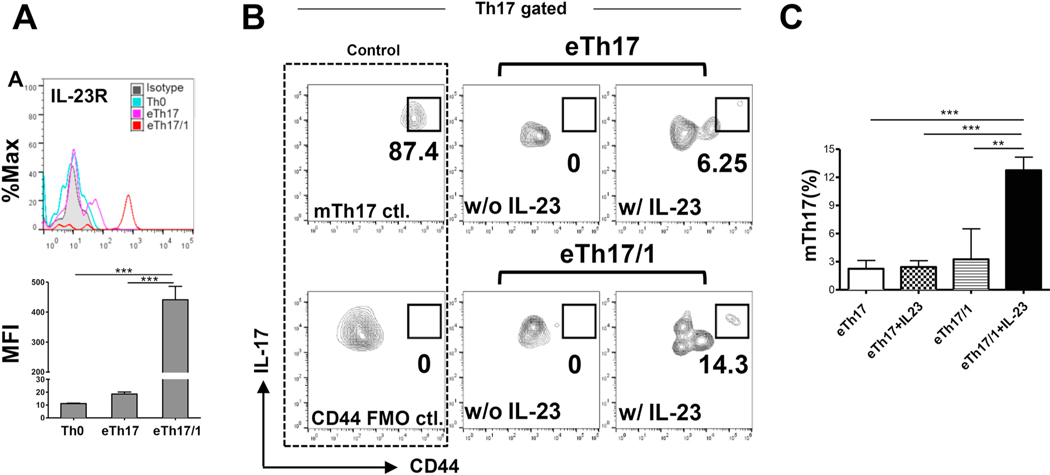
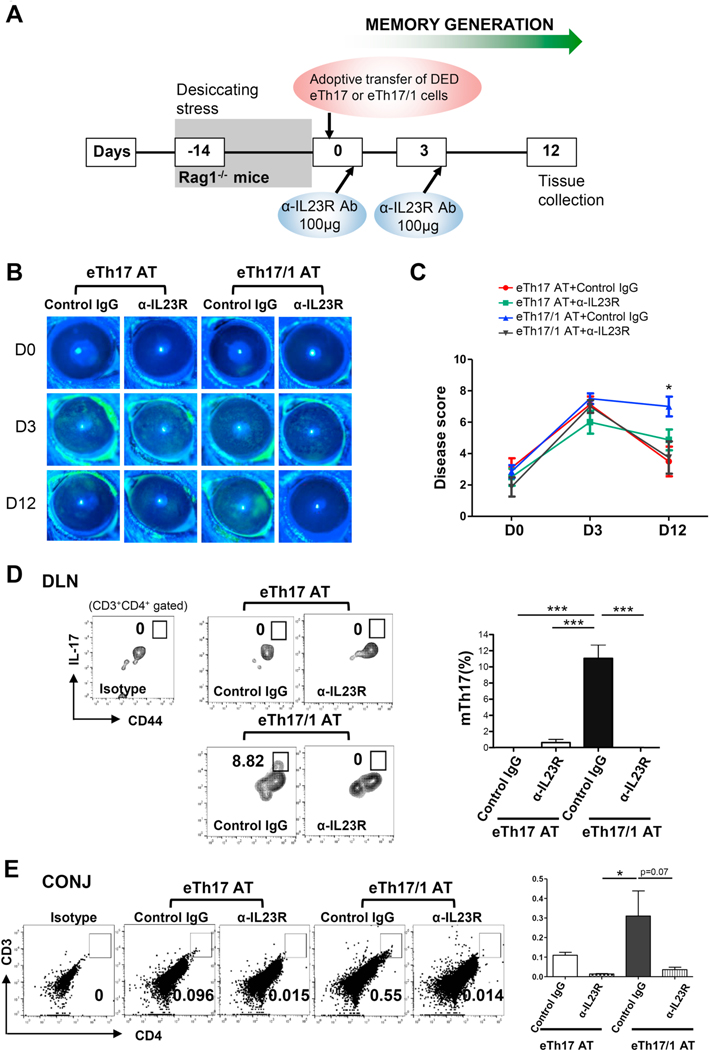
Effector Th17 cells, including IFN-γ-IL-17+ (eTh17) and IFN-γ+IL-17+ (eTh17/1) subsets, play critical pathogenic functions in the induction of autoimmunity. As acute inflammation subsides, a small proportion of the effectors survive and convert to memory Th17 cells (mTh17), which sustain chronic inflammation in autoimmune diseases. Herein, we investigated the differential contributions of eTh17 versus eTh17/1 to the memory pool using an experimental model of ocular autoimmune disease. Our results show that adoptive transfer of Tbx21-/- CD4+ T cells or conditional deletion of Tbx21 in Th17 cells leads to diminished eTh17/1 in acute phase and functionally compromised mTh17 in chronic phase. Further, adoptive transfer of disease-specific eTh17/1, but not eTh17, leads to generation of mTh17 and sustained ocular inflammation. Collectively, our data demonstrate that T-bet-dependent eTh17/1 cells generated during the acute inflammation are the principal effector precursors of pathogenic mTh17 cells that sustain the chronicity of autoimmune inflammation.
Keywords: Effector Th17/1; Memory Th17; T-bet.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
