

Specific inflammatory osteoclast precursors induced during chronic inflammation give rise to highly active osteoclasts associated with inflammatory bone loss
- PMID: 35396510
- PMCID: PMC8993801
- DOI: 10.1038/s41413-022-00206-z
Specific inflammatory osteoclast precursors induced during chronic inflammation give rise to highly active osteoclasts associated with inflammatory bone loss
Abstract
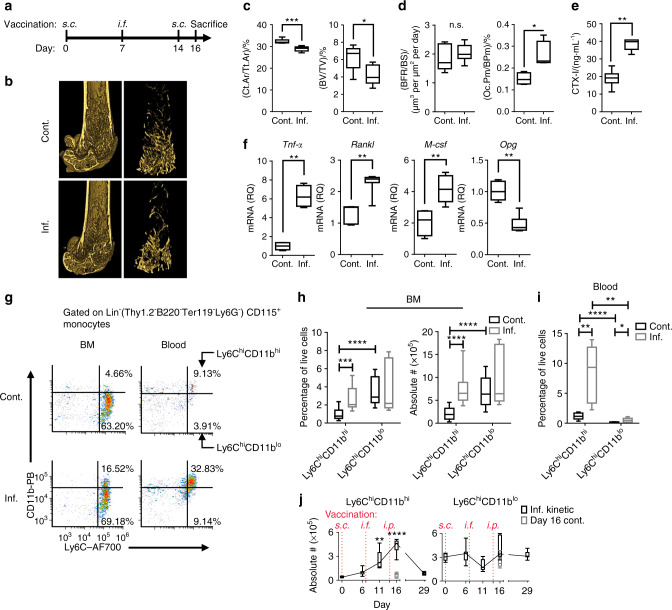
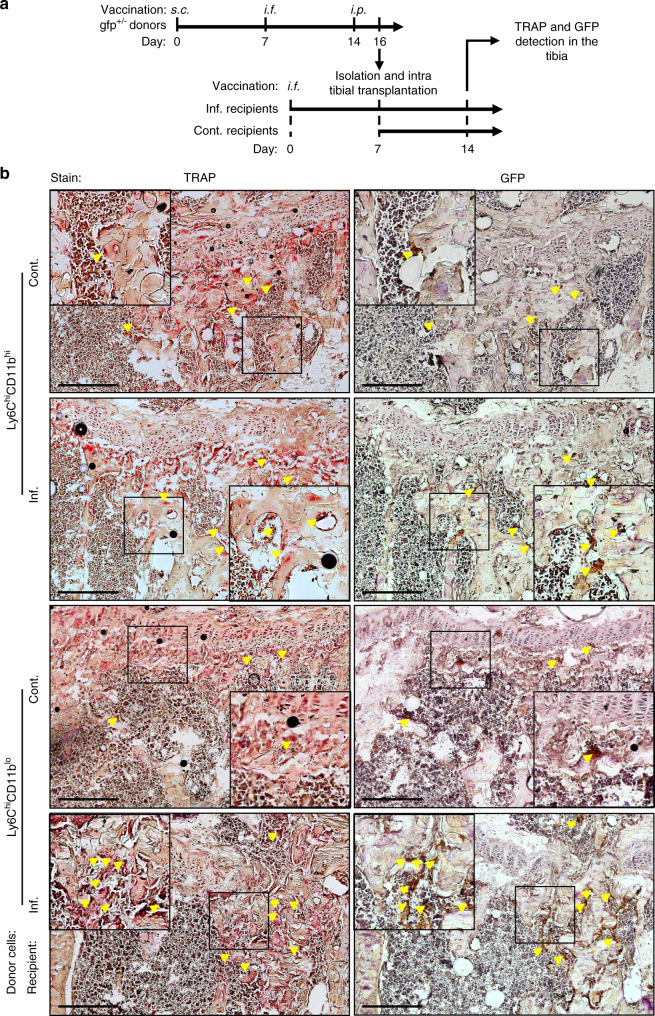
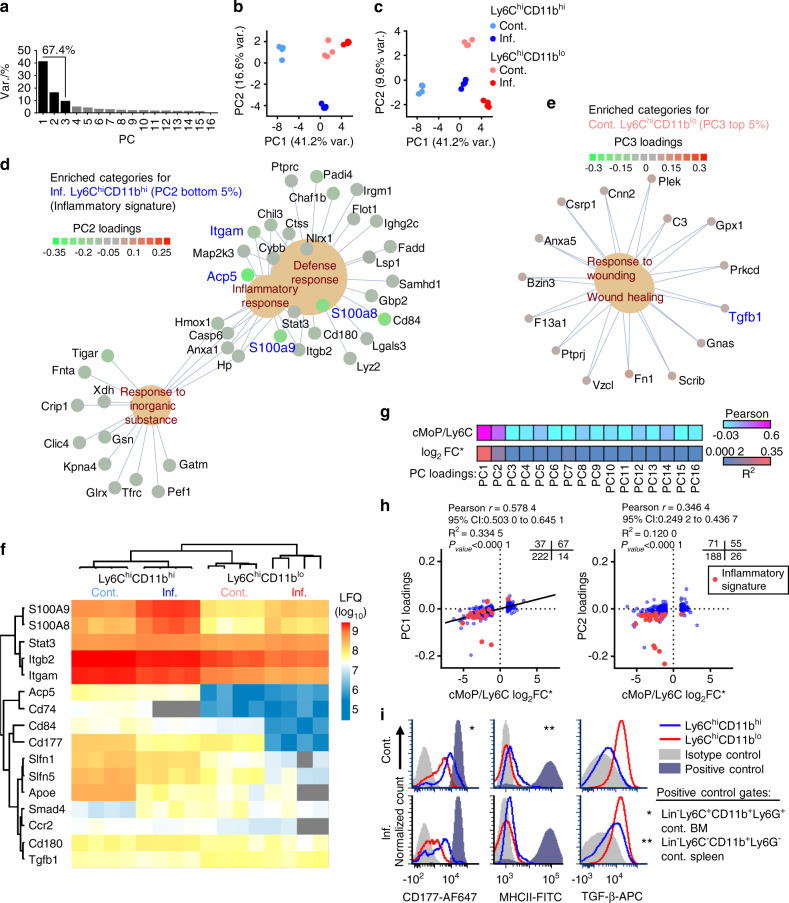
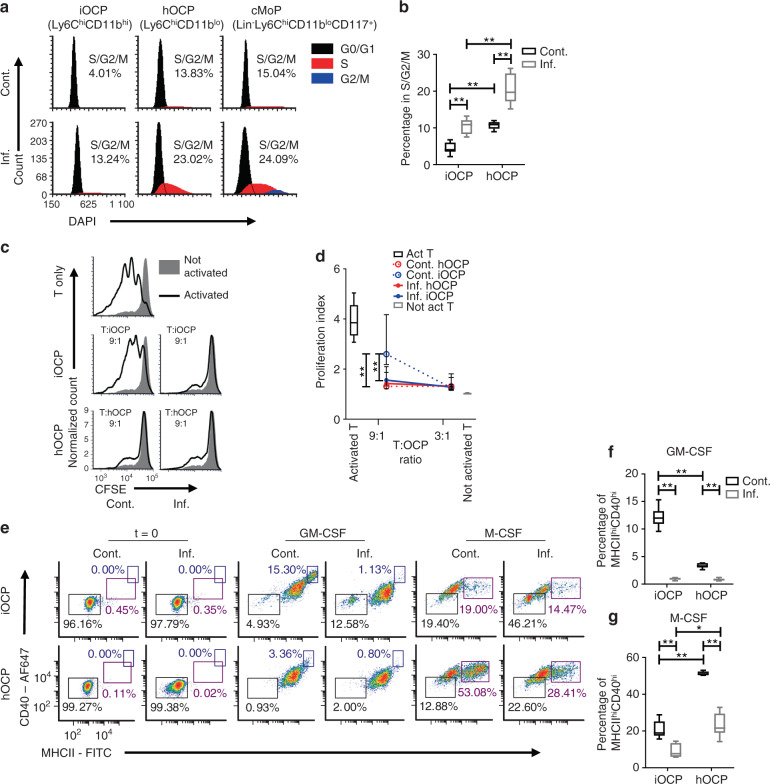
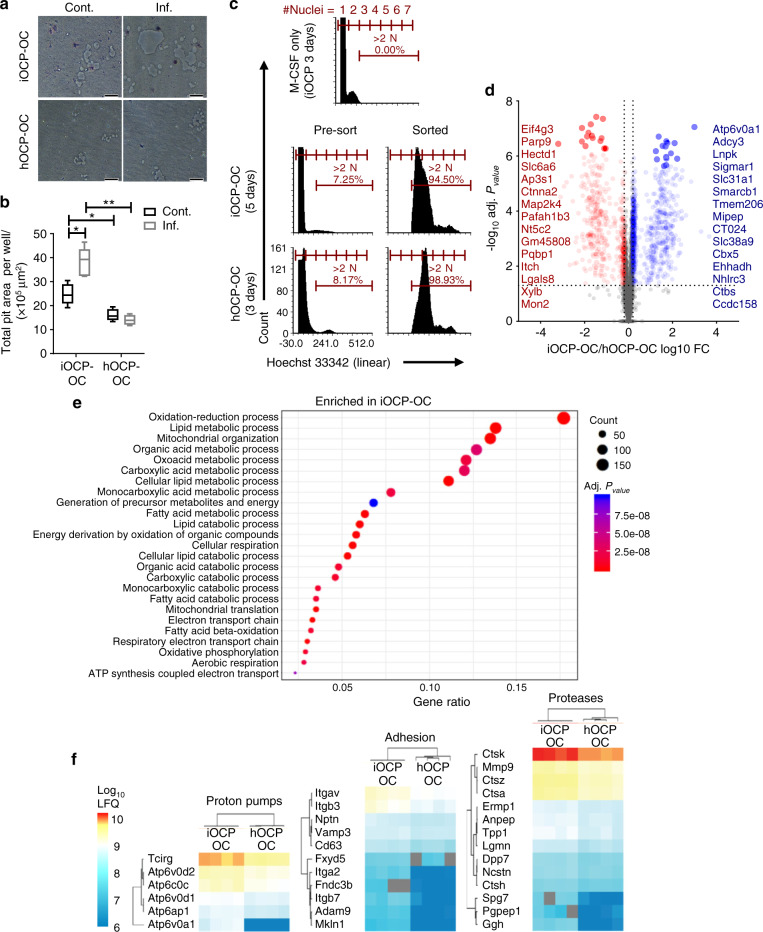
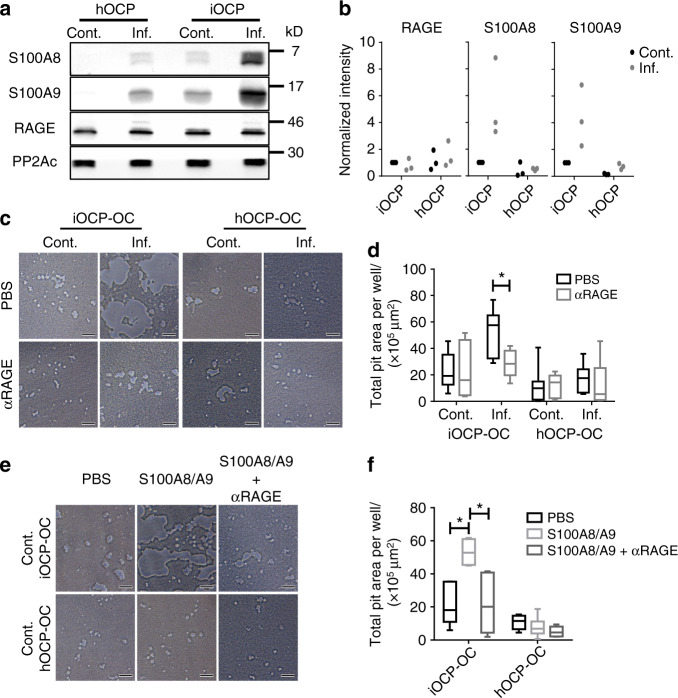
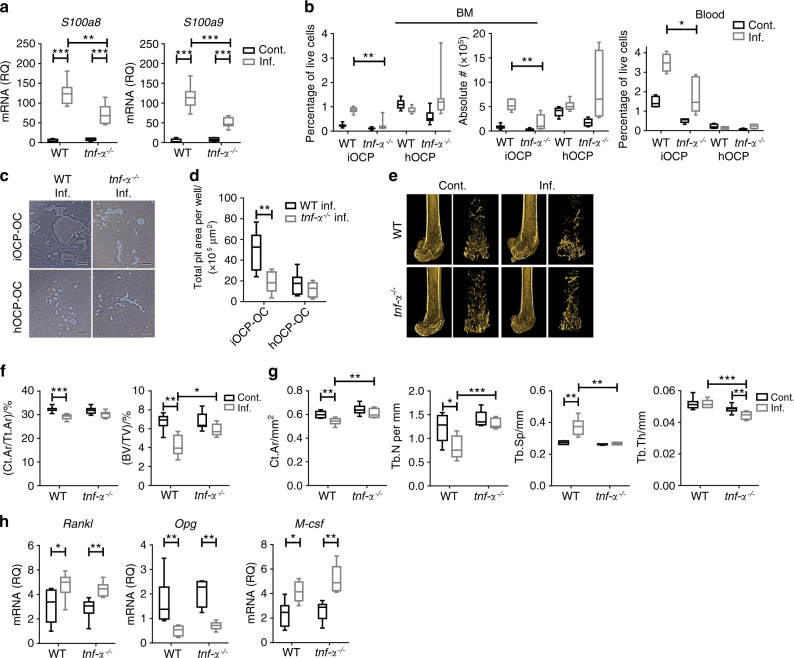
Elevated osteoclast (OC) activity is a major contributor to inflammatory bone loss (IBL) during chronic inflammatory diseases. However, the specific OC precursors (OCPs) responding to inflammatory cues and the underlying mechanisms leading to IBL are poorly understood. We identified two distinct OCP subsets: Ly6ChiCD11bhi inflammatory OCPs (iOCPs) induced during chronic inflammation, and homeostatic Ly6ChiCD11blo OCPs (hOCPs) which remained unchanged. Functional and proteomic characterization revealed that while iOCPs were rare and displayed low osteoclastogenic potential under normal conditions, they expanded during chronic inflammation and generated OCs with enhanced activity. In contrast, hOCPs were abundant and manifested high osteoclastogenic potential under normal conditions but generated OCs with low activity and were unresponsive to the inflammatory environment. Osteoclasts derived from iOCPs expressed higher levels of resorptive and metabolic proteins than those generated from hOCPs, highlighting that different osteoclast populations are formed by distinct precursors. We further identified the TNF-α and S100A8/A9 proteins as key regulators that control the iOCP response during chronic inflammation. Furthermore, we demonstrated that the response of iOCPs but not that of hOCPs was abrogated in tnf-α-/- mice, in correlation with attenuated IBL. Our findings suggest a central role for iOCPs in IBL induction. iOCPs can serve as potential biomarkers for IBL detection and possibly as new therapeutic targets to combat IBL in a wide range of inflammatory conditions.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
LinkOut - more resources
Full Text Sources
Research Materials
