

Dynamics of absolute and relative disparity processing in human visual cortex
- PMID: 35398280
- PMCID: PMC9205266
- DOI: 10.1016/j.neuroimage.2022.119186
Dynamics of absolute and relative disparity processing in human visual cortex
Abstract
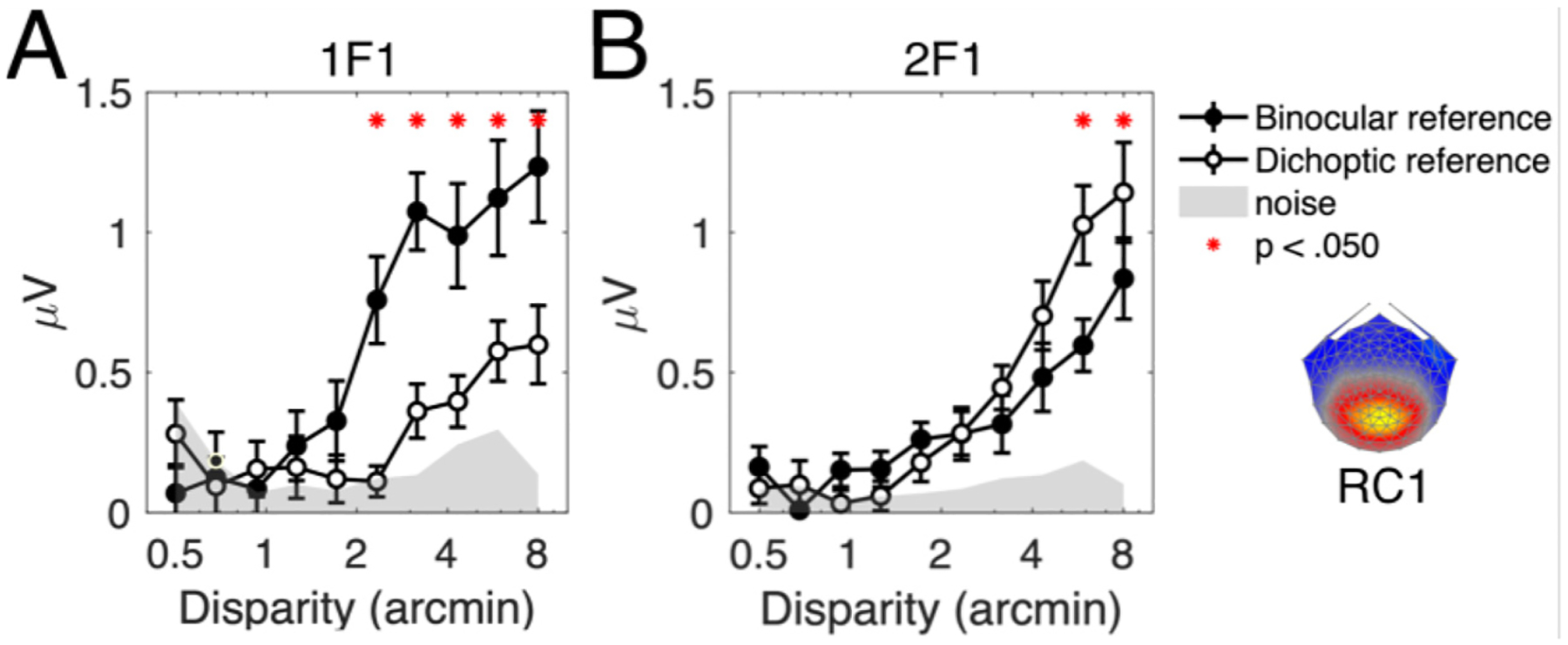
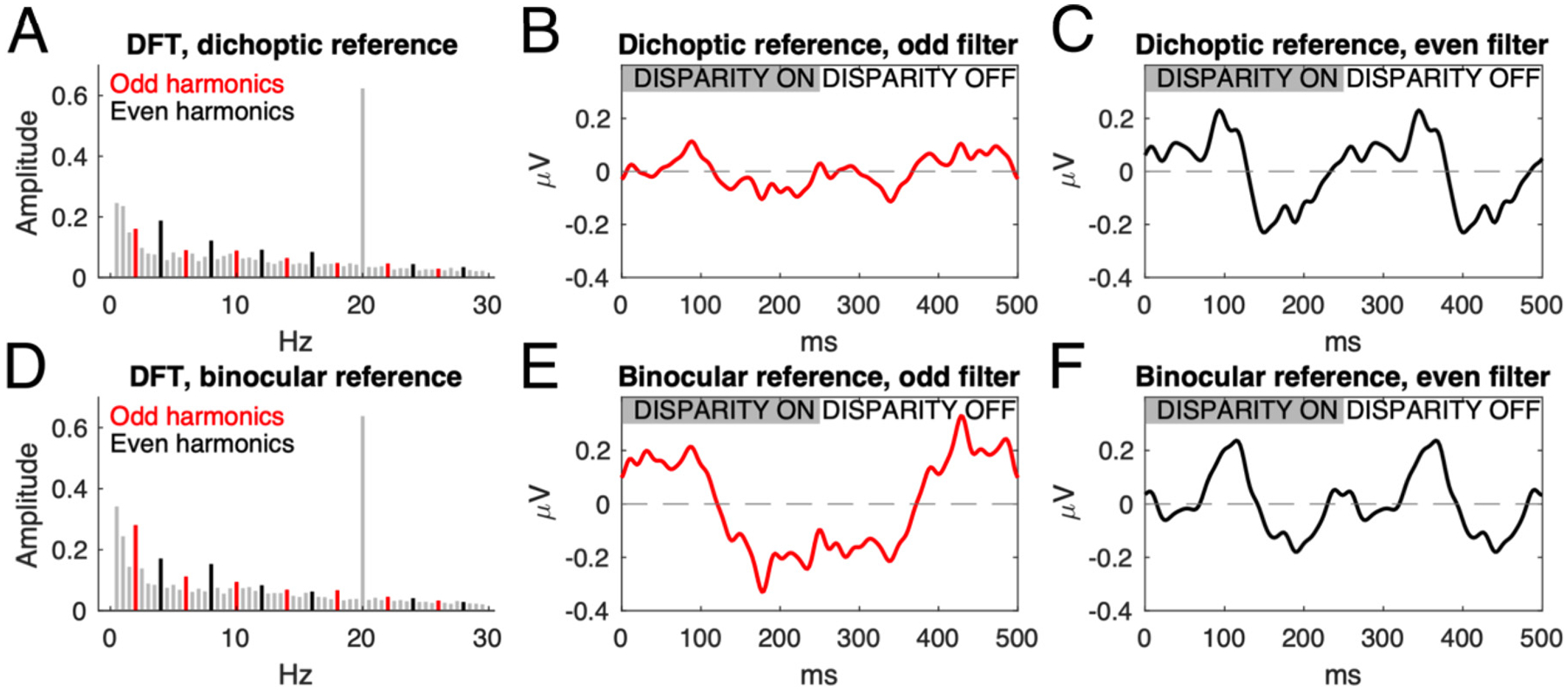
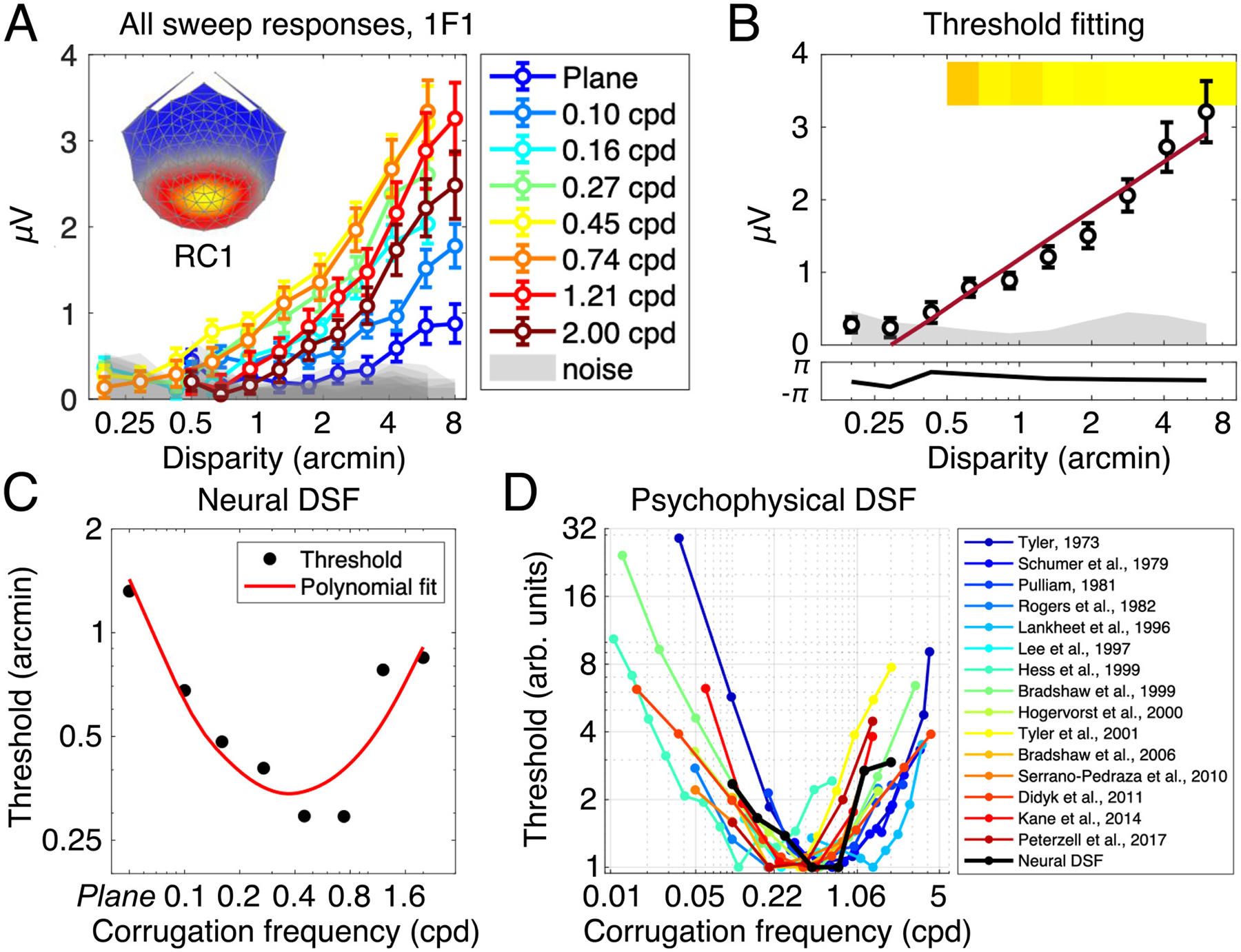
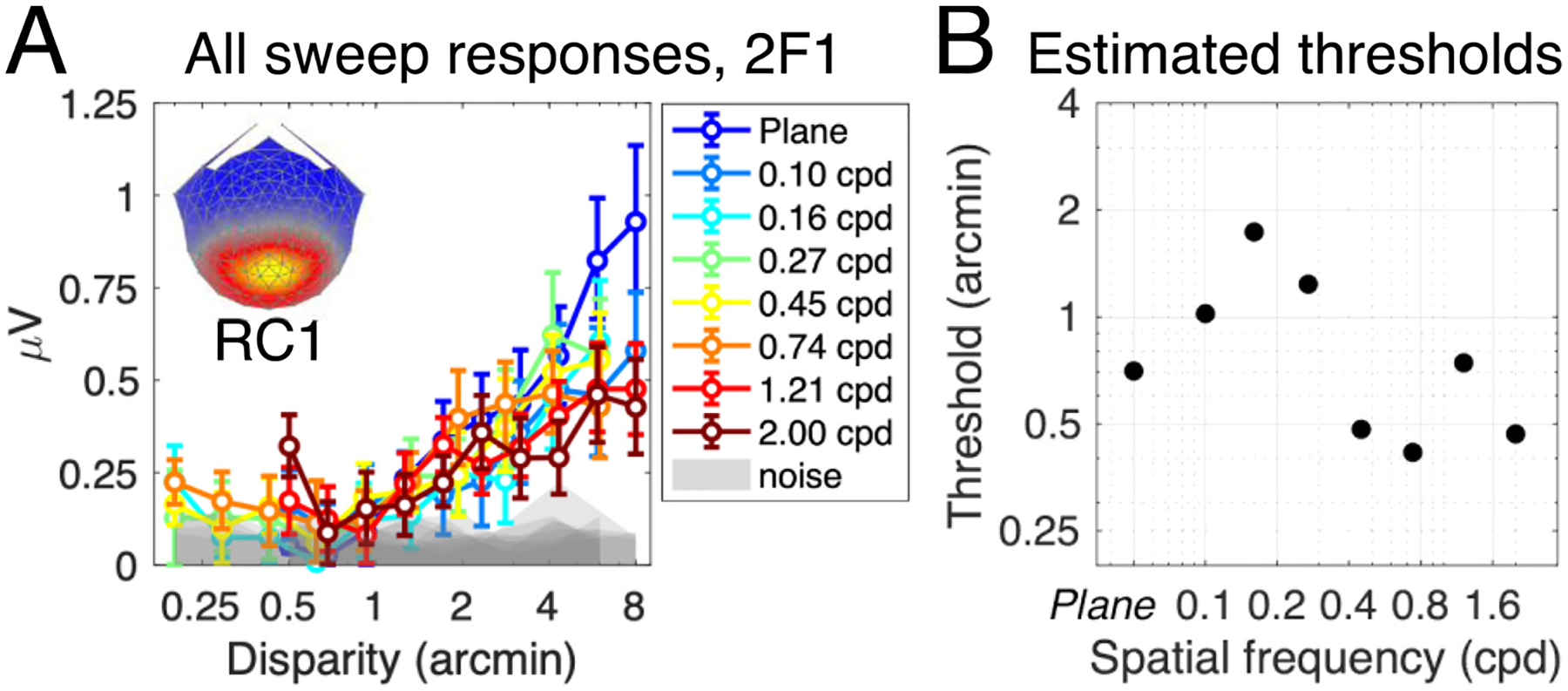
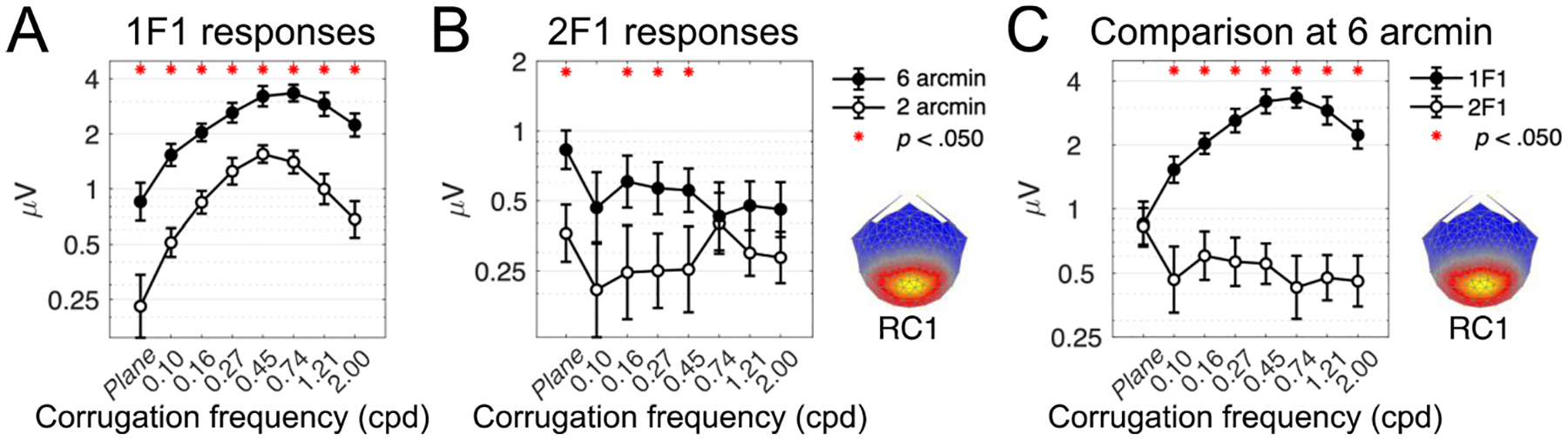
Cortical processing of binocular disparity is believed to begin in V1 where cells are sensitive to absolute disparity, followed by the extraction of relative disparity in higher visual areas. While much is known about the cortical distribution and spatial tuning of disparity-selective neurons, the relationship between their spatial and temporal properties is less well understood. Here, we use steady-state Visual Evoked Potentials and dynamic random dot stereograms to characterize the temporal dynamics of spatial mechanisms in human visual cortex that are primarily sensitive to either absolute or relative disparity. Stereograms alternated between disparate and non-disparate states at 2 Hz. By varying the disparity-defined spatial frequency content of the stereograms from a planar surface to corrugated ones, we biased responses towards absolute vs. relative disparities. Reliable Components Analysis was used to derive two dominant sources from the 128 channel EEG records. The first component (RC1) was maximal over the occipital pole. In RC1, first harmonic responses were sustained, tuned for corrugation frequency, and sensitive to the presence of disparity references, consistent with prior psychophysical sensitivity measurements. By contrast, the second harmonic, associated with transient processing, was not spatially tuned and was indifferent to references, consistent with it being generated by an absolute disparity mechanism. Thus, our results reveal a duplex coding strategy in the disparity domain, where relative disparities are computed via sustained mechanisms and absolute disparities are computed via transient mechanisms.
Keywords: Binocular vision; Disparity; SSVEP; Sustained; Transient.
Copyright © 2022. Published by Elsevier Inc.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
