

Impact of Dietary Palmitic Acid on Lipid Metabolism
- PMID: 35399673
- PMCID: PMC8983927
- DOI: 10.3389/fnut.2022.861664
Impact of Dietary Palmitic Acid on Lipid Metabolism
Abstract
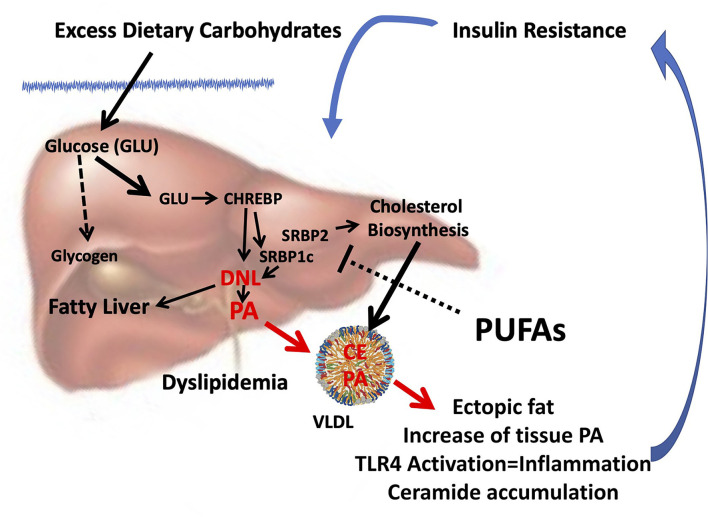
Palmitic acid (PA) is ubiquitously present in dietary fat guaranteeing an average intake of about 20 g/d. The relative high requirement and relative content in the human body, which accounts for 20-30% of total fatty acids (FAs), is justified by its relevant nutritional role. In particular physiological conditions, such as in the fetal stage or in the developing brain, the respectively inefficient placental and brain blood-barrier transfer of PA strongly induces its endogenous biosynthesis from glucose via de novo lipogenesis (DNL) to secure a tight homeostatic control of PA tissue concentration required to exert its multiple physiological activities. However, pathophysiological conditions (insulin resistance) are characterized by a sustained DNL in the liver and aimed at preventing the excess accumulation of glucose, which result in increased tissue content of PA and disrupted homeostatic control of its tissue concentration. This leads to an overaccumulation of tissue PA, which results in dyslipidemia, increased ectopic fat accumulation, and inflammatory tone via toll-like receptor 4. Any change in dietary saturated FAs (SFAs) usually reflects a complementary change in polyunsaturated FA (PUFA) intake. Since PUFA particularly n-3 highly PUFA, suppress lipogenic gene expression, their reduction in intake rather than excess of dietary SFA may promote endogenous PA production via DNL. Thereby, the increase in tissue PA and its deleterious consequences from dysregulated DNL can be mistakenly attributed to dietary intake of PA.
Keywords: de novo lipogenesis; dietary fatty acids; fatty acid metabolism; palmitic acid; saturated/unsaturated ratio.
Copyright © 2022 Murru, Manca, Carta and Banni.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous