

Restricted Proliferation During Neurogenesis Contributes to Regionalisation of the Amphioxus Nervous System
- PMID: 35401089
- PMCID: PMC8987370
- DOI: 10.3389/fnins.2022.812223
Restricted Proliferation During Neurogenesis Contributes to Regionalisation of the Amphioxus Nervous System
Abstract
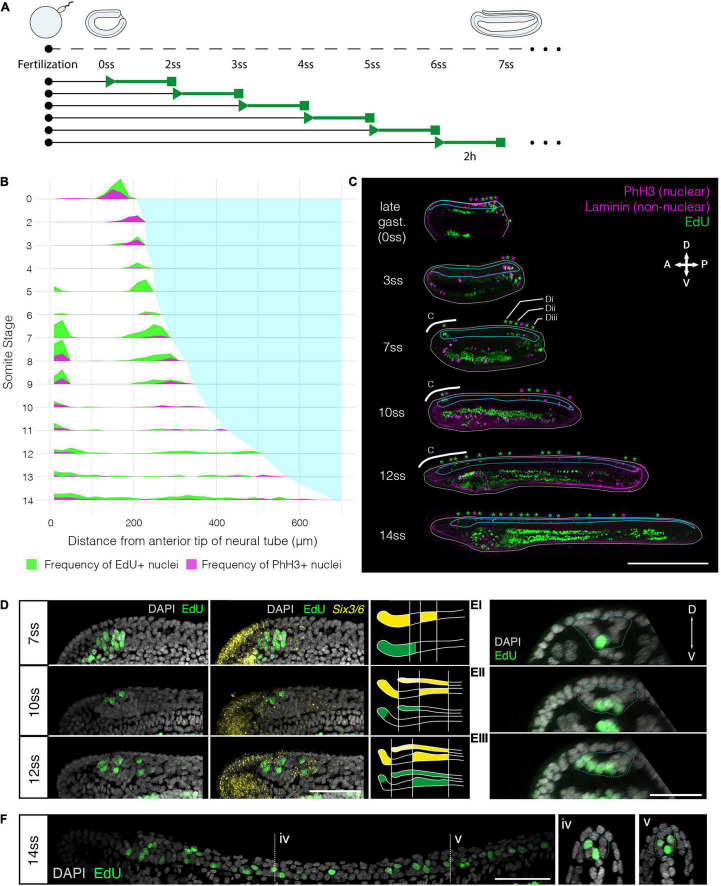
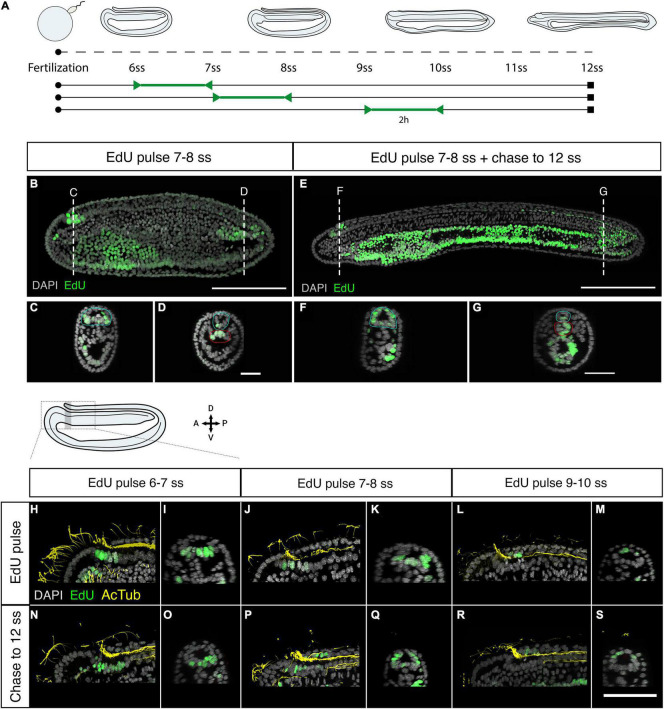
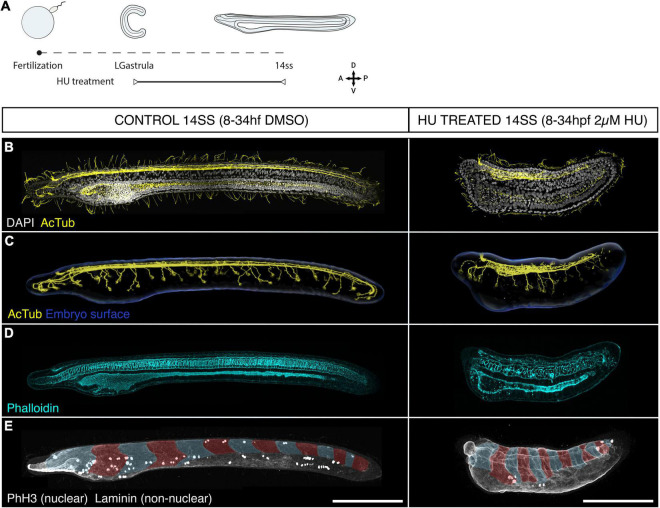
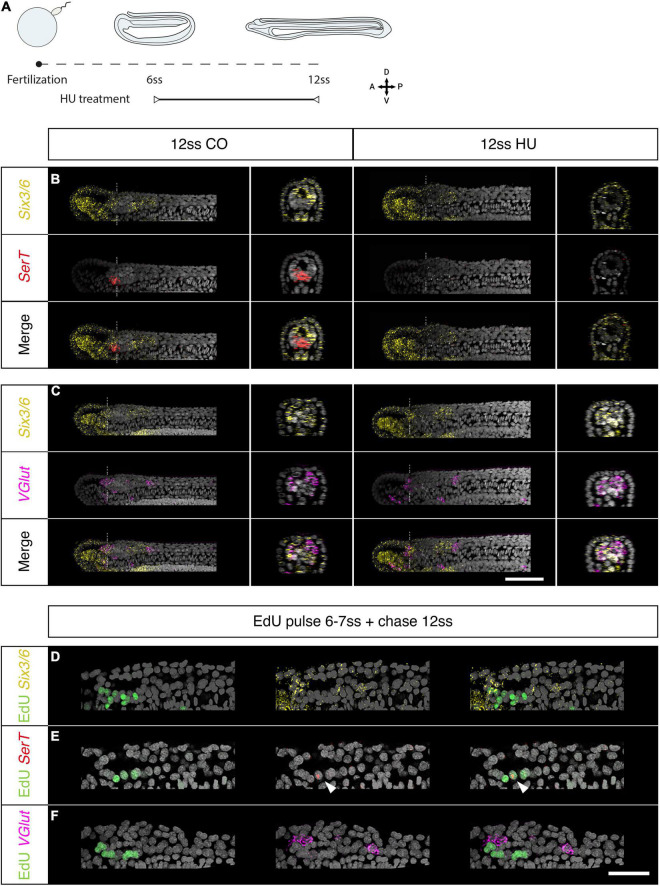
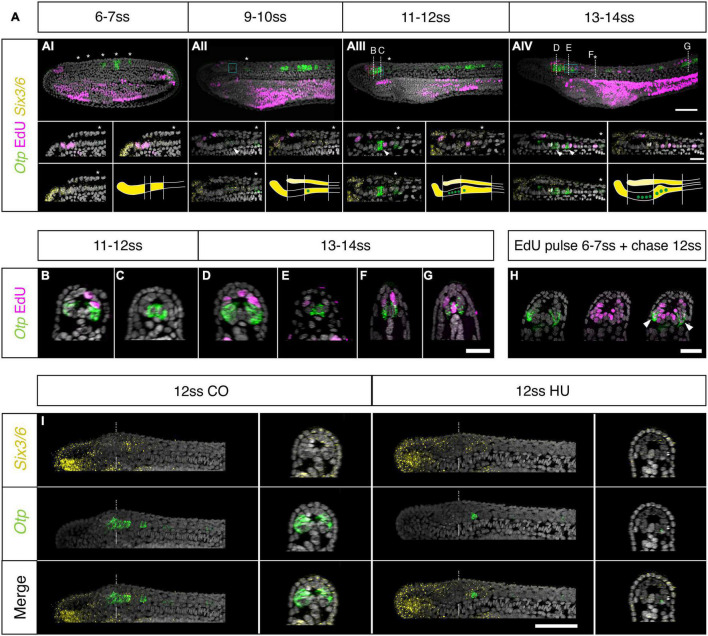
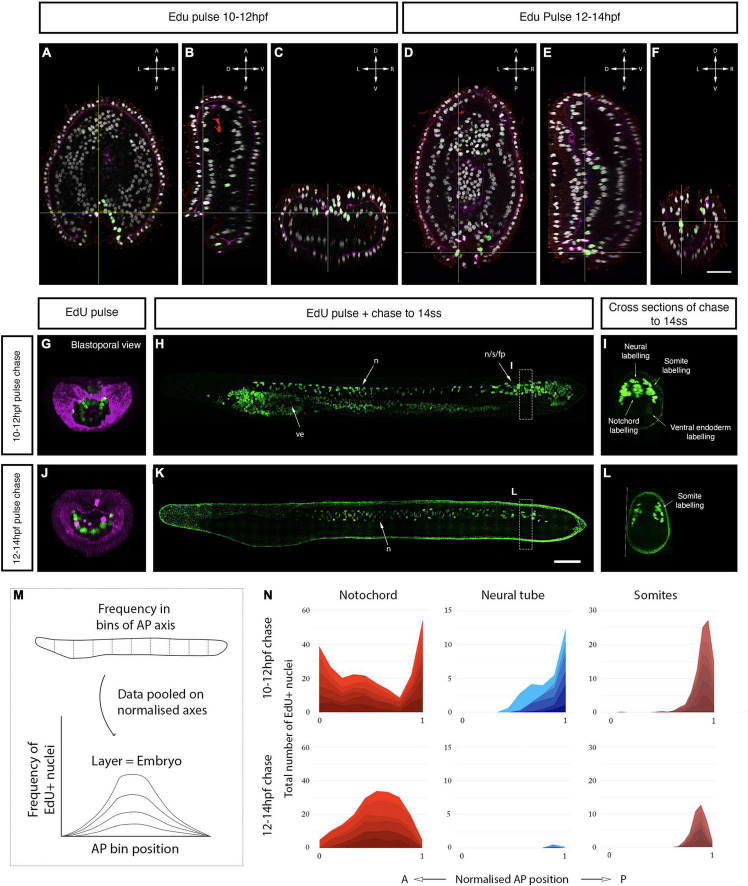
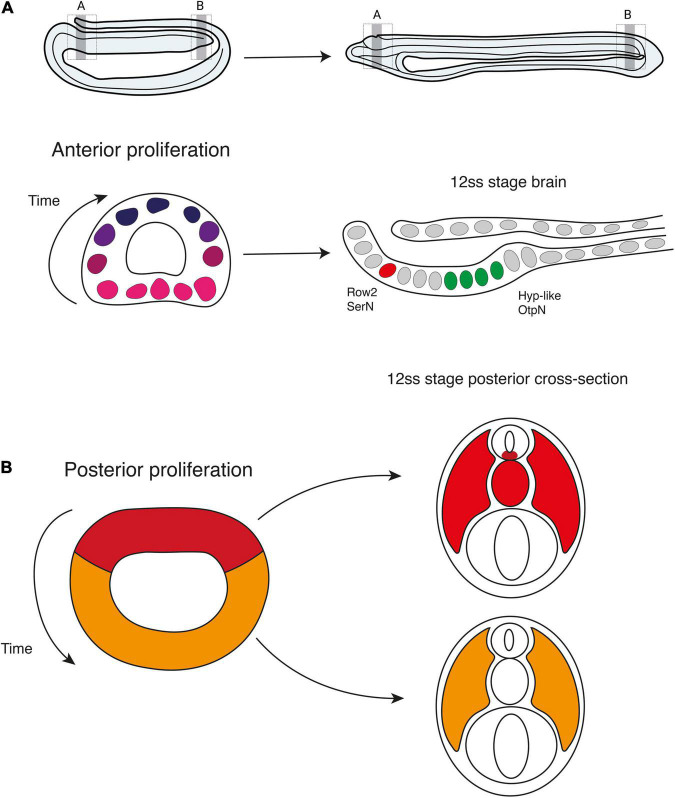
The central nervous system of the cephalochordate amphioxus consists of a dorsal neural tube with an anterior brain. Two decades of gene expression analyses in developing amphioxus embryos have shown that, despite apparent morphological simplicity, the amphioxus neural tube is highly regionalised at the molecular level. However, little is known about the morphogenetic mechanisms regulating the spatiotemporal emergence of cell types at distinct sites of the neural axis and how their arrangements contribute to the overall neural architecture. In vertebrates, proliferation is key to provide appropriate cell numbers of specific types to particular areas of the nervous system as development proceeds, but in amphioxus proliferation has never been studied at this level of detail, nor in the specific context of neurogenesis. Here, we describe the dynamics of cell division during the formation of the central nervous system in amphioxus embryos, and identify specific regions of the nervous system that depend on proliferation of neuronal precursors at precise time-points for their maturation. By labelling proliferating cells in vivo at specific time points in development, and inhibiting cell division during neurulation, we demonstrate that localised proliferation in the anterior cerebral vesicle is required to establish the full cell type repertoire of the frontal eye complex and the putative hypothalamic region of the amphioxus brain, while posterior proliferating progenitors, which were found here to derive from the dorsal lip of the blastopore, contribute to elongation of the caudal floor plate. Between these proliferative domains, we find that trunk nervous system differentiation is independent from cell division, in which proliferation decreases during neurulation and resumes at the early larval stage. Taken together, our results highlight the importance of proliferation as a tightly controlled mechanism for shaping and regionalising the amphioxus neural axis during development, by addition of new cells fated to particular types, or by influencing tissue geometry.
Keywords: EdU pulse-chase; HCR; amphioxus; axial progenitors; brain development; chordate evolution; neurogenesis; proliferation.
Copyright © 2022 Gattoni, Andrews and Benito-Gutiérrez.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Retinoic acid signaling and neurogenic niche regulation in the developing peripheral nervous system of the cephalochordate amphioxus.Cell Mol Life Sci. 2018 Jul;75(13):2407-2429. doi: 10.1007/s00018-017-2734-3. Epub 2018 Jan 31. Cell Mol Life Sci. 2018. PMID: 29387904 Free PMC article.
-
Characterisation of amphioxus HNF-3 genes: conserved expression in the notochord and floor plate.Dev Biol. 1997 Mar 1;183(1):74-85. doi: 10.1006/dbio.1996.8481. Dev Biol. 1997. PMID: 9119116
-
Sequence and developmental expression of AmphiDll, an amphioxus Distal-less gene transcribed in the ectoderm, epidermis and nervous system: insights into evolution of craniate forebrain and neural crest.Development. 1996 Sep;122(9):2911-20. doi: 10.1242/dev.122.9.2911. Development. 1996. PMID: 8787764
-
Origin and evolution of the chordate central nervous system: insights from amphioxus genoarchitecture.Int J Dev Biol. 2017;61(10-11-12):655-664. doi: 10.1387/ijdb.170258jg. Int J Dev Biol. 2017. PMID: 29319114 Review.
-
Sensory systems in amphioxus: a window on the ancestral chordate condition.Brain Behav Evol. 2004;64(3):148-62. doi: 10.1159/000079744. Brain Behav Evol. 2004. PMID: 15353907 Review.
Cited by
-
The Expression of Shmt Genes in Amphioxus Suggests a Role in Tissue Proliferation Rather than in Neurotransmission.Cells. 2025 Jul 13;14(14):1071. doi: 10.3390/cells14141071. Cells. 2025. PMID: 40710324 Free PMC article.
-
An ancient apical patterning system sets the position of the forebrain in chordates.Sci Adv. 2025 Jan 24;11(4):eadq4731. doi: 10.1126/sciadv.adq4731. Epub 2025 Jan 24. Sci Adv. 2025. PMID: 39854450 Free PMC article.
-
A feather star is born: embryonic development and nervous system organization in the crinoid Antedon mediterranea.Open Biol. 2024 Aug;14(8):240115. doi: 10.1098/rsob.240115. Epub 2024 Aug 21. Open Biol. 2024. PMID: 39165121 Free PMC article.
-
Evolutionary origin of the chordate nervous system revealed by amphioxus developmental trajectories.Nat Ecol Evol. 2024 Sep;8(9):1693-1710. doi: 10.1038/s41559-024-02469-7. Epub 2024 Jul 18. Nat Ecol Evol. 2024. PMID: 39025981
References
-
- Albuixech-Crespo B., López-Blanch L., Burguera D., Maeso I., Sánchez-Arrones L., Moreno-Bravo J. A., et al. (2017). Molecular regionalization of the developing amphioxus neural tube challenges major partitions of the vertebrate. brain. PLoS Biol. 15:e2001573. 10.1371/journal.pbio.2001573 - DOI - PMC - PubMed
-
- Andrews T. G., Gattoni G., Busby L., Schwimmer M. A., Benito-Gutiérrez È. (2020). “Hybridization chain reaction for quantitative and multiplex imaging of gene expression in amphioxus embryos and adult tissues,” in In Situ Hybridization Protocols, eds Nielsen B. S., Jones J. (Berlin: Springer Nature; ), 179–194. 10.1007/978-1-0716-0623-0 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources