

Dual RNA Sequencing Reveals the Genome-Wide Expression Profiles During the Compatible and Incompatible Interactions Between Solanum tuberosum and Phytophthora infestans
- PMID: 35401650
- PMCID: PMC8993506
- DOI: 10.3389/fpls.2022.817199
Dual RNA Sequencing Reveals the Genome-Wide Expression Profiles During the Compatible and Incompatible Interactions Between Solanum tuberosum and Phytophthora infestans
Abstract
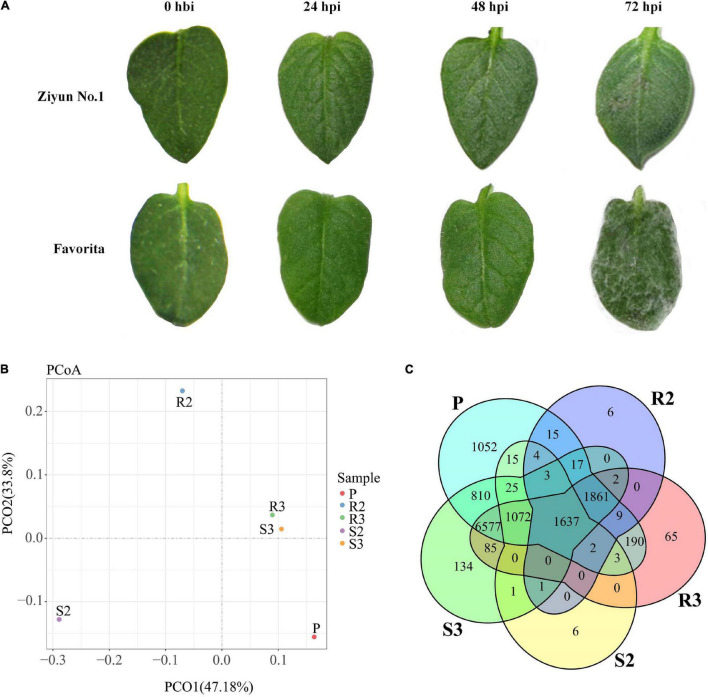
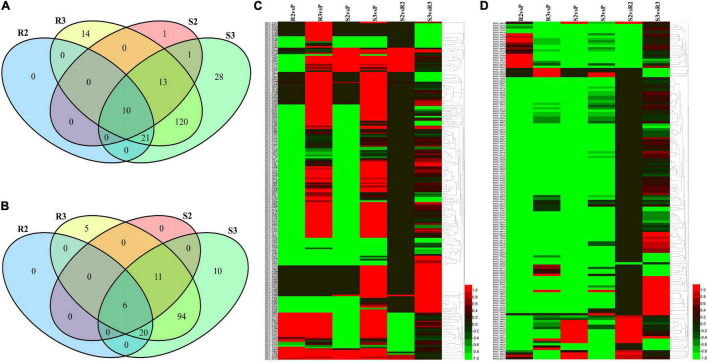
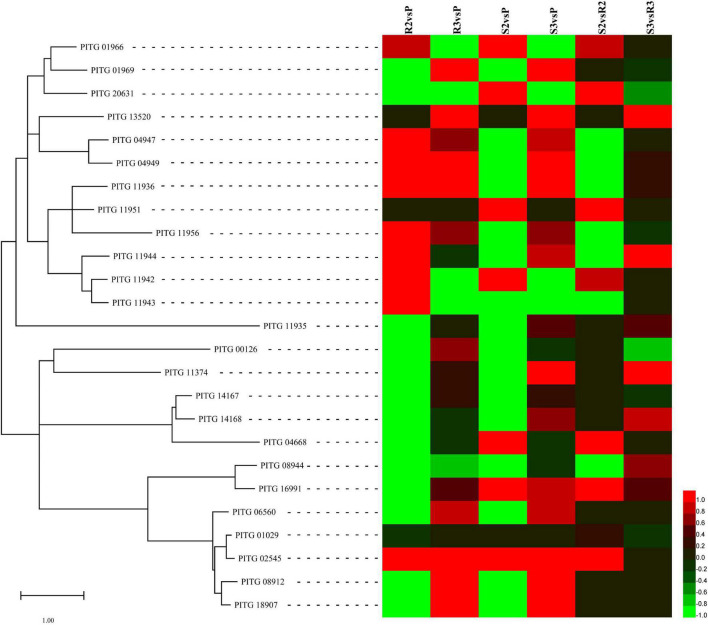
Late blight, caused by Phytophthora infestans (P. infestans), is a devastating plant disease. P. infestans genome encodes hundreds of effectors, complicating the interaction between the pathogen and its host and making it difficult to understand the interaction mechanisms. In this study, the late blight-resistant potato cultivar Ziyun No.1 and the susceptible potato cultivar Favorita were infected with P. infestans isolate SCPZ16-3-1 to investigate the global expression profiles during the compatible and incompatible interactions using dual RNA sequencing (RNA-seq). Most of the expressed Arg-X-Leu-Arg (RXLR) effector genes were suppressed during the first 24 h of infection, but upregulated after 24 h. Moreover, P. infestans induced more specifically expressed genes (SEGs), including RXLR effectors and cell wall-degrading enzymes (CWDEs)-encoding genes, in the compatible interaction. The resistant potato activated a set of biotic stimulus responses and phenylpropanoid biosynthesis SEGs, including kirola-like protein, nucleotide-binding site-leucine-rich repeat (NBS-LRR), disease resistance, and kinase genes. Conversely, the susceptible potato cultivar upregulated more kinase, pathogenesis-related genes than the resistant cultivar. This study is the first study to characterize the compatible and incompatible interactions between P. infestans and different potato cultivars and provides the genome-wide expression profiles for RXLR effector, CWDEs, NBS-LRR protein, and kinase-encoding genes.
Keywords: Phytophthora infestans; compatible interaction; different potato cultivars; dual RNA-seq; incompatible interaction.
Copyright © 2022 Li, Hu, Fan, Chen, Jiang, Huang and Tao.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
-
- AC’t Hoen P., Ariyurek Y., Thygesen H. H., Vreugdenhil E., Vossen R. H., De Menezes R. X., et al. (2008). Deep sequencing-based expression analysis shows major advances in robustness, resolution and inter-lab portability over five microarray platforms. Nucleic Acids Res. 36:e141. 10.1093/nar/gkn705 - DOI - PMC - PubMed
-
- Armstrong M. R., Whisson S. C., Pritchard L., Bos J. I. B., Venter E., Avrova A. O., et al. (2005). An ancestral oomycete locus contains late blight avirulence gene Avr3a, encoding a protein that is recognized in the host cytoplasm. Proc. Natl. Acad. Sci. U.S.A. 102 7766–7771. 10.1073/pnas.0500113102 - DOI - PMC - PubMed
-
- Ballvora A., Ercolano M. R., Weiß J., Meksem K., Bormann C. A., Oberhagemann P., et al. (2002). The R1 gene for potato resistance to late blight (Phytophthora infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant J. 30 361–371. 10.1046/j.1365-313x.2001.01292.x - DOI - PubMed
-
- Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc.: Ser. B (Methodological) 57 289–300.
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous