

Persistence of Virus-Specific Antibody after Depletion of Memory B Cells
- PMID: 35404084
- PMCID: PMC9093124
- DOI: 10.1128/jvi.00026-22
Persistence of Virus-Specific Antibody after Depletion of Memory B Cells
Abstract
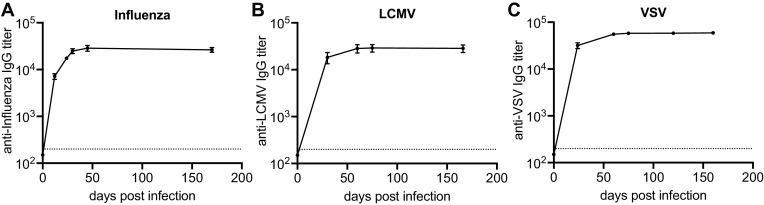
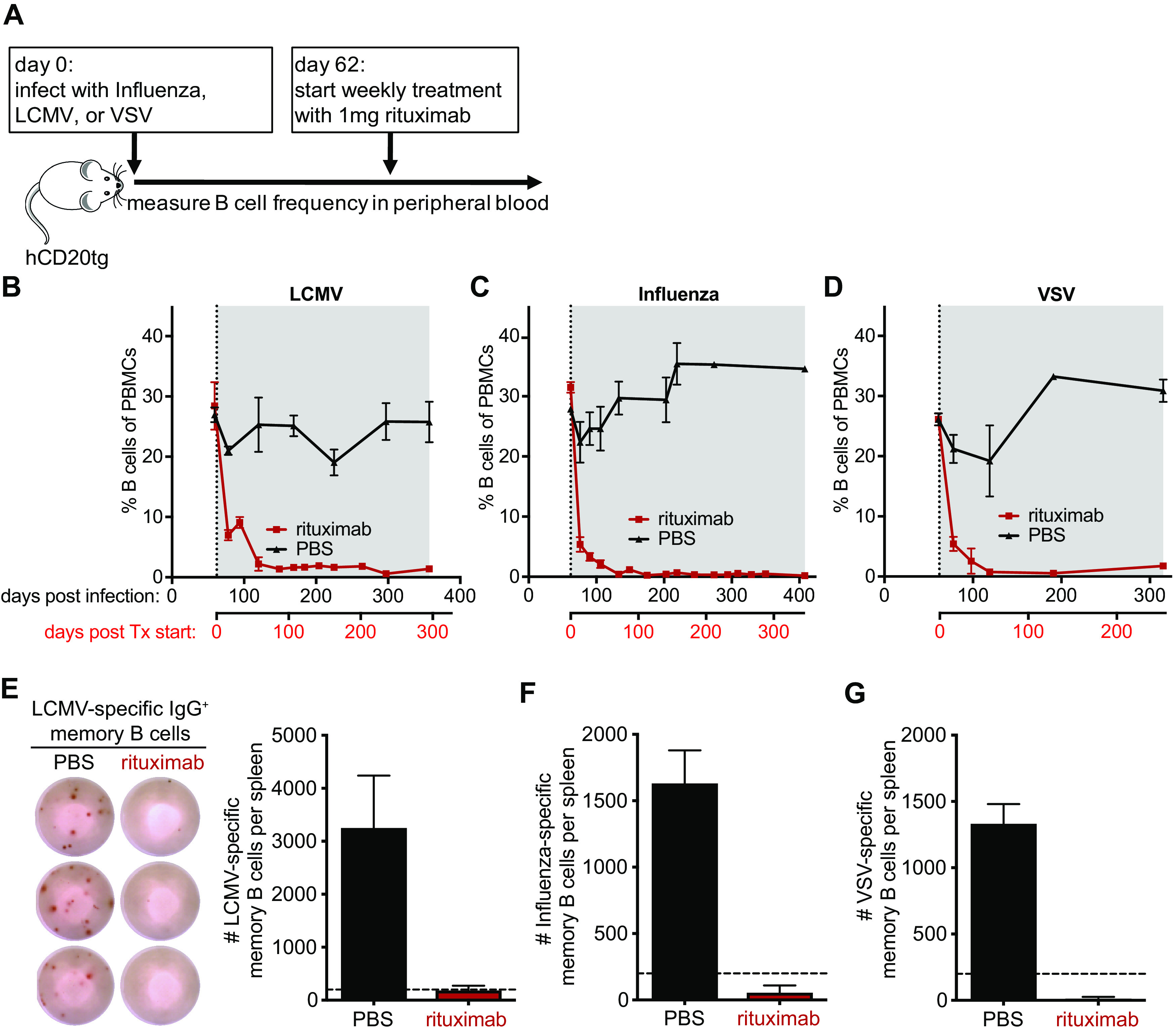
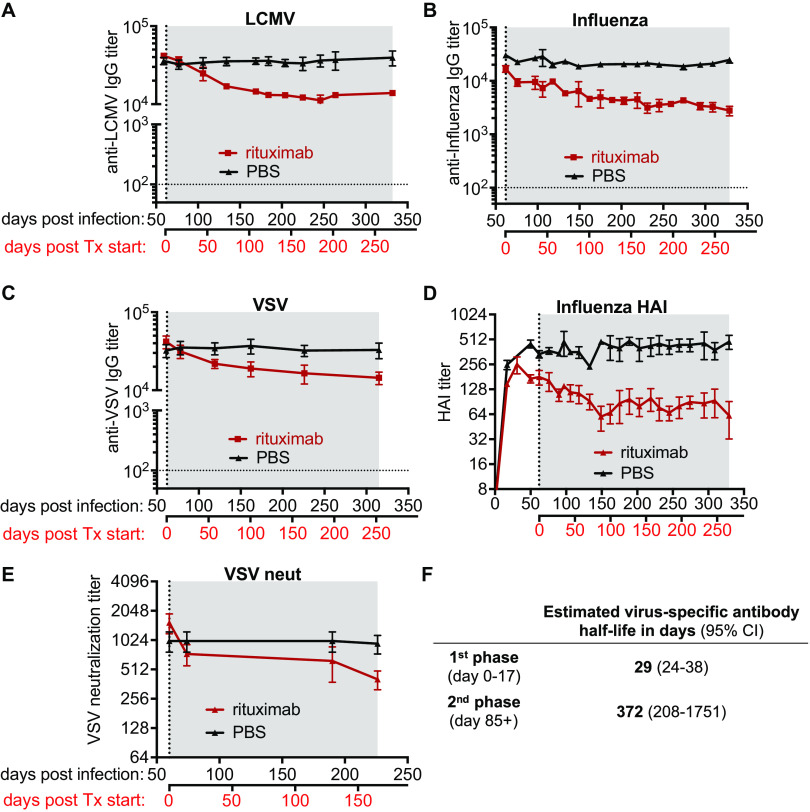
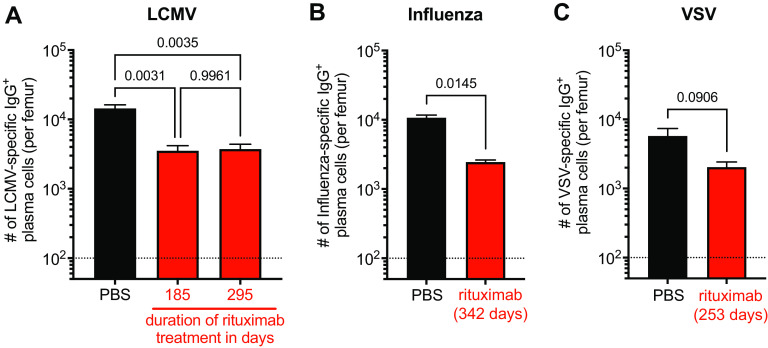
Humoral immunity is a major component of the adaptive immune response against viruses and other pathogens with pathogen-specific antibody acting as the first line of defense against infection. Virus-specific antibody levels are maintained by continual secretion of antibody by plasma cells residing in the bone marrow. This raises the important question of how the virus-specific plasma cell population is stably maintained and whether memory B cells are required to replenish plasma cells, balancing their loss arising from their intrinsic death rate. In this study, we examined the longevity of virus-specific antibody responses in the serum of mice following acute viral infection with three different viruses: lymphocytic choriomeningitis virus (LCMV), influenza virus, and vesicular stomatitis virus (VSV). To investigate the contribution of memory B cells to the maintenance of virus-specific antibody levels, we employed human CD20 transgenic mice, which allow for the efficient depletion of B cells with rituximab, a human CD20-specific monoclonal antibody. Mice that had resolved an acute infection with LCMV, influenza virus, or VSV were treated with rituximab starting at 2 months after infection, and the treatment was continued for up to a year postinfection. This treatment regimen with rituximab resulted in efficient depletion of B cells (>95%), with virus-specific memory B cells being undetectable. There was an early transient drop in the antibody levels after rituximab treatment followed by a plateauing of the curve with virus-specific antibody levels remaining relatively stable (half-life of 372 days) for up to a year after infection in the absence of memory B cells. The number of virus-specific plasma cells in the bone marrow were consistent with the changes seen in serum antibody levels. Overall, our data show that virus-specific plasma cells in the bone marrow are intrinsically long-lived and can maintain serum antibody titers for extended periods of time without requiring significant replenishment from memory B cells. These results provide insight into plasma cell longevity and have implications for B cell depletion regimens in cancer and autoimmune patients in the context of vaccination in general and especially for COVID-19 vaccines. IMPORTANCE Following vaccination or primary virus infection, virus-specific antibodies provide the first line of defense against reinfection. Plasma cells residing in the bone marrow constitutively secrete antibodies, are long-lived, and can thus maintain serum antibody levels over extended periods of time in the absence of antigen. Our data, in the murine model system, show that virus-specific plasma cells are intrinsically long-lived but that some reseeding by memory B cells might occur. Our findings demonstrate that, due to the longevity of plasma cells, virus-specific antibody levels remain relatively stable in the absence of memory B cells and have implications for vaccination.
Keywords: B cell responses; antibodies; immune memory; plasma cells; viral immunity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
