

CCR2/CCR5 inhibitor permits the radiation-induced effector T cell infiltration in pancreatic adenocarcinoma
- PMID: 35404390
- PMCID: PMC9006312
- DOI: 10.1084/jem.20211631
CCR2/CCR5 inhibitor permits the radiation-induced effector T cell infiltration in pancreatic adenocarcinoma
Abstract
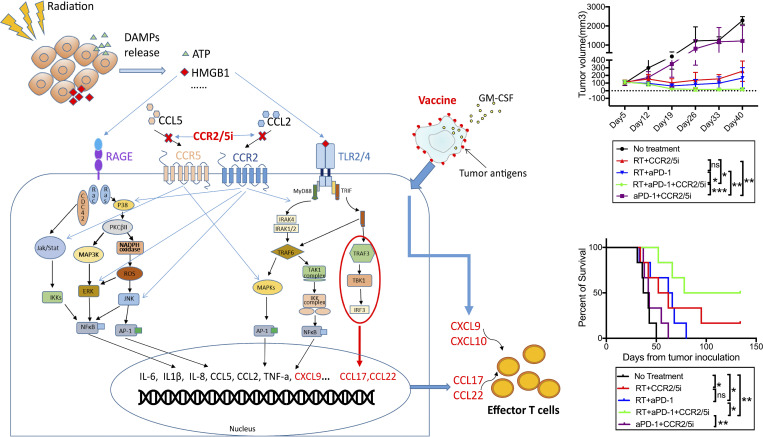
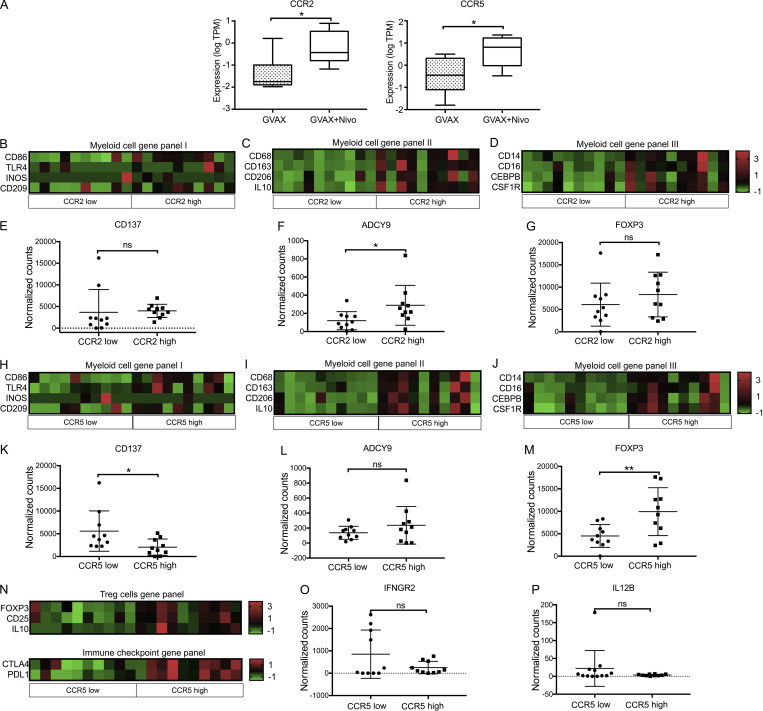
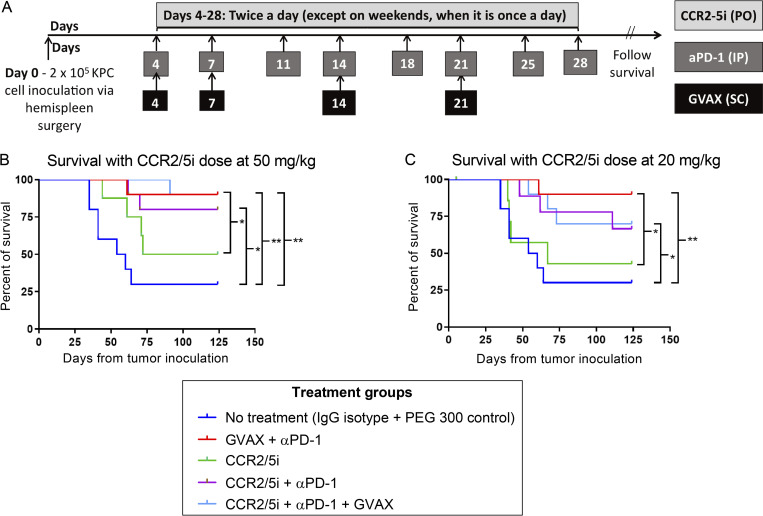
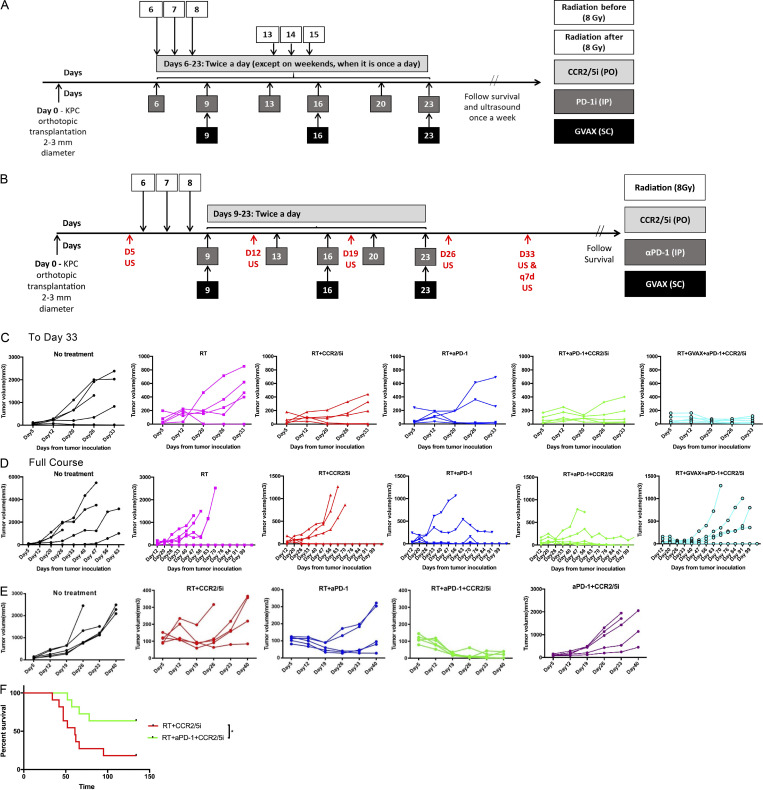
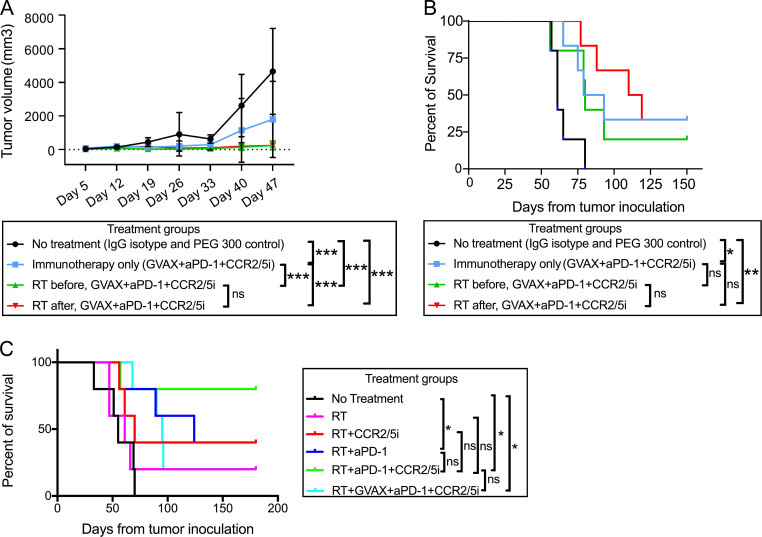
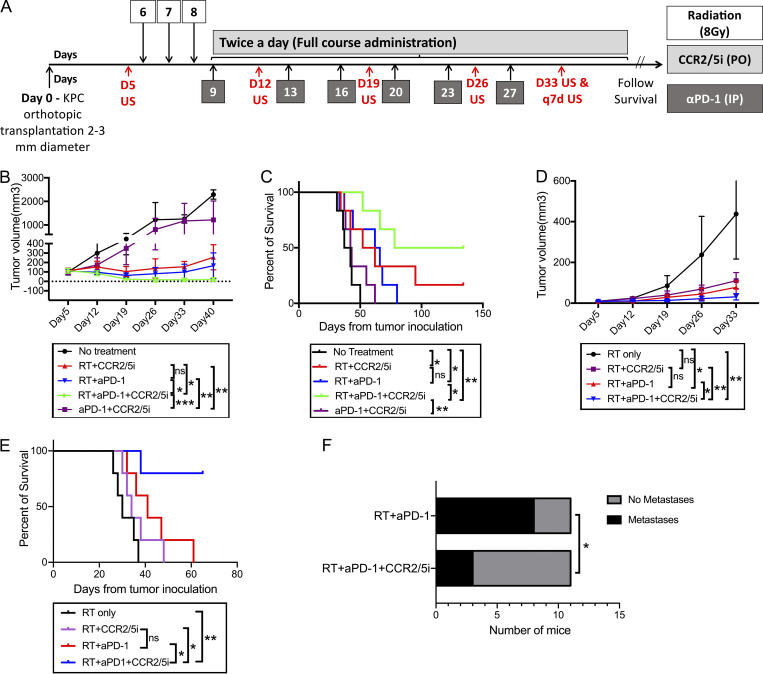
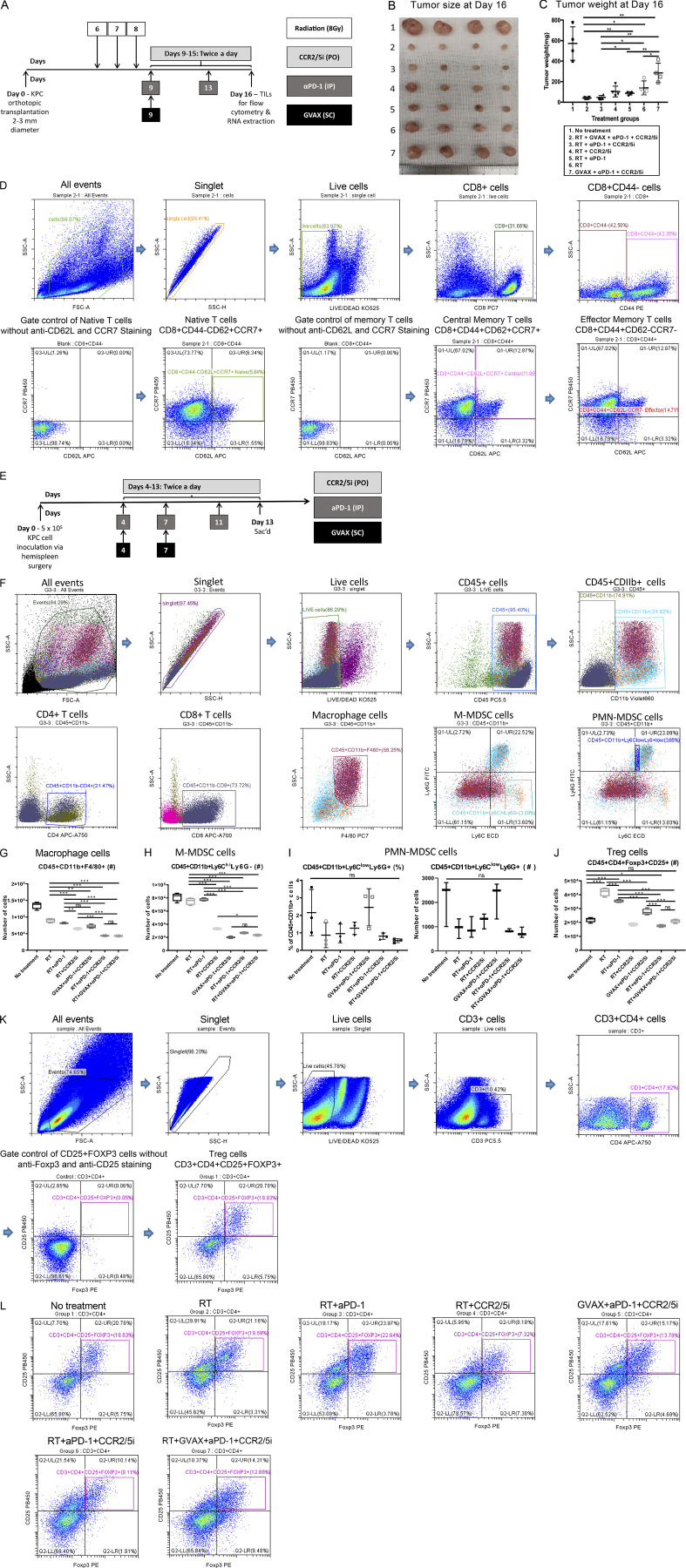
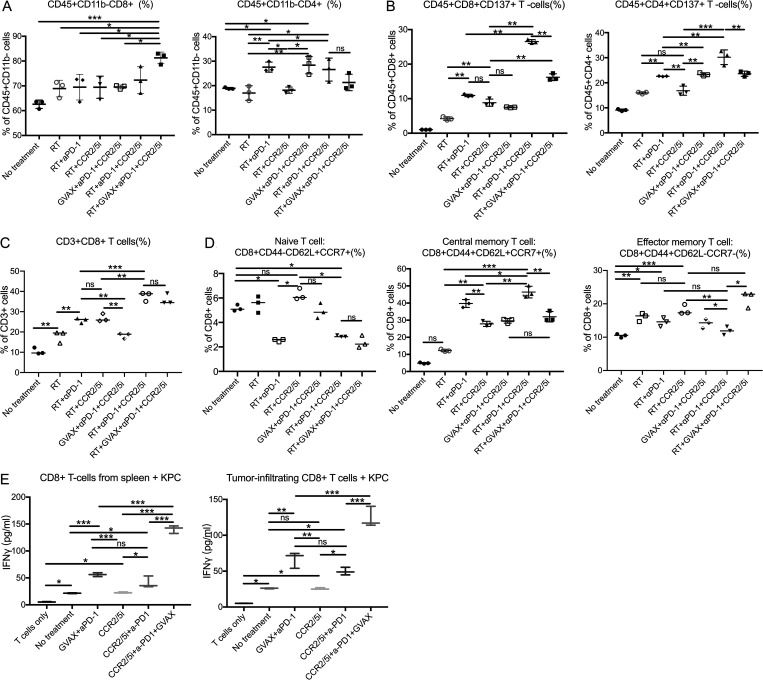
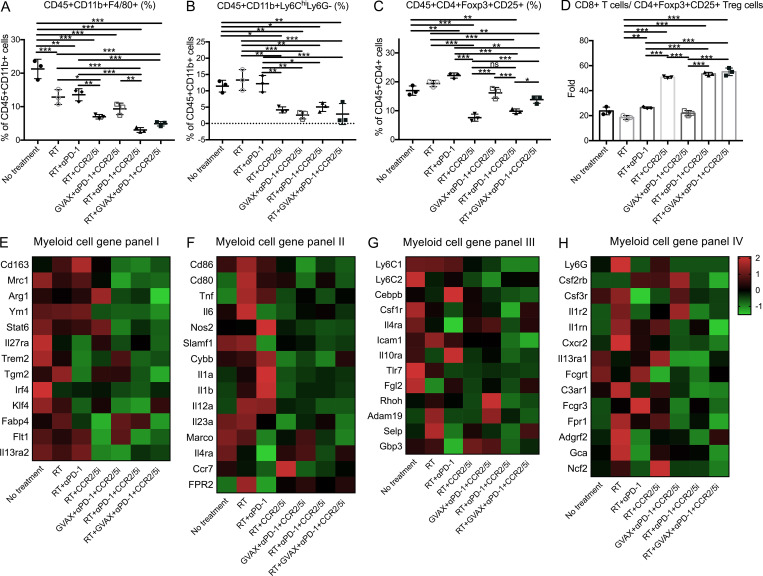
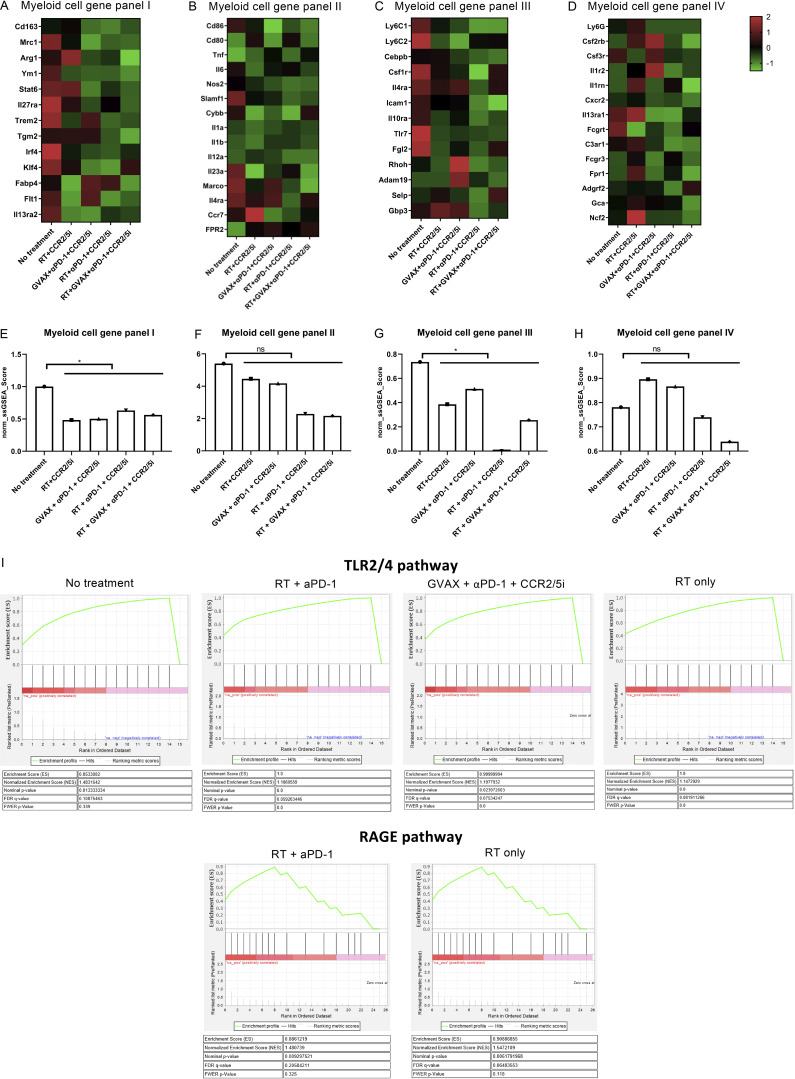
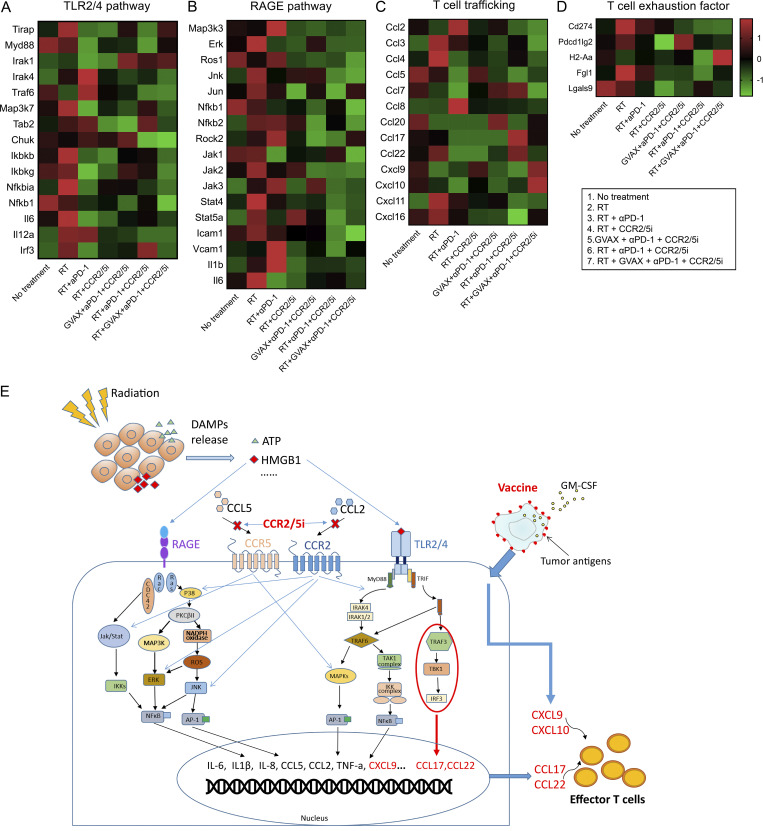
The resistance of pancreatic ductal adenocarcinoma (PDAC) to immune checkpoint inhibitors (ICIs) is attributed to the immune-quiescent and -suppressive tumor microenvironment (TME). We recently found that CCR2 and CCR5 were induced in PDAC following treatment with anti-PD-1 antibody (αPD-1); thus, we examined PDAC vaccine or radiation therapy (RT) as T cell priming mechanisms together with BMS-687681, a dual antagonist of CCR2 and CCR5 (CCR2/5i), in combination with αPD-1 as new treatment strategies. Using PDAC mouse models, we demonstrated that RT followed by αPD-1 and prolonged treatment with CCR2/5i conferred better antitumor efficacy than other combination treatments tested. The combination of RT + αPD-1 + CCR2/5i enhanced intratumoral effector and memory T cell infiltration but suppressed regulatory T cell, M2-like tumor-associated macrophage, and myeloid-derived suppressive cell infiltration. RNA sequencing showed that CCR2/5i partially inhibited RT-induced TLR2/4 and RAGE signaling, leading to decreased expression of immunosuppressive cytokines including CCL2/CCL5, but increased expression of effector T cell chemokines such as CCL17/CCL22. This study thus supports the clinical development of CCR2/5i in combination with RT and ICIs for PDAC treatment.
© 2022 Wang et al.
Conflict of interest statement
Disclosures: K. Fujiwara reported “other” from The Beacon Group, personal fees from Medcase, and grants from Japan Society for the Promotion of Science outside the submitted work. C. Twyman-Saint Victor reported personal fees from Bristol Myers Squibb during the conduct of the study and personal fees from Bristol Myers Squibb outside the submitted work. L. Zheng reported grants from Bristol-Myers Squibb, Merck, Astrazeneca, iTeos, Amgen, NovaRock, Inxmed, and Halozyme, and personal fees from Biosion, Alphamab, NovaRock, Ambrx, Xilio, QED, Natera, Tempus, Pfizer, Johnson and Johnson, Novagenesis, Snow Lake Capitals, BioArdis, and Mingruizhiyao during the conduct of the study. In addition, L. Zheng holds shares at Alphamab and Mingruizhiyao. No other disclosures were reported.
Figures

Similar articles
-
Targeting both tumour-associated CXCR2+ neutrophils and CCR2+ macrophages disrupts myeloid recruitment and improves chemotherapeutic responses in pancreatic ductal adenocarcinoma.Gut. 2018 Jun;67(6):1112-1123. doi: 10.1136/gutjnl-2017-313738. Epub 2017 Dec 1. Gut. 2018. PMID: 29196437 Free PMC article.
-
Targeting myeloid-inflamed tumor with anti-CSF-1R antibody expands CD137+ effector T-cells in the murine model of pancreatic cancer.J Immunother Cancer. 2018 Nov 13;6(1):118. doi: 10.1186/s40425-018-0435-6. J Immunother Cancer. 2018. PMID: 30424804 Free PMC article.
-
Interrogating the immune-modulating roles of radiation therapy for a rational combination with immune-checkpoint inhibitors in treating pancreatic cancer.J Immunother Cancer. 2020 Jul;8(2):e000351. doi: 10.1136/jitc-2019-000351. J Immunother Cancer. 2020. PMID: 32675194 Free PMC article.
-
Immune Checkpoint Inhibition for Pancreatic Ductal Adenocarcinoma: Current Limitations and Future Options.Front Immunol. 2018 Aug 15;9:1878. doi: 10.3389/fimmu.2018.01878. eCollection 2018. Front Immunol. 2018. PMID: 30158932 Free PMC article. Review.
-
Current advances and outlooks in immunotherapy for pancreatic ductal adenocarcinoma.Mol Cancer. 2020 Feb 15;19(1):32. doi: 10.1186/s12943-020-01151-3. Mol Cancer. 2020. PMID: 32061257 Free PMC article. Review.
Cited by
-
Therapeutic Perspectives of HIV-Associated Chemokine Receptor (CCR5 and CXCR4) Antagonists in Carcinomas.Int J Mol Sci. 2022 Dec 28;24(1):478. doi: 10.3390/ijms24010478. Int J Mol Sci. 2022. PMID: 36613922 Free PMC article. Review.
-
Transcriptional control of pancreatic cancer immunosuppression by metabolic enzyme CD73 in a tumor-autonomous and -autocrine manner.Nat Commun. 2023 Jun 8;14(1):3364. doi: 10.1038/s41467-023-38578-3. Nat Commun. 2023. PMID: 37291128 Free PMC article.
-
Consensus, debate, and prospective on pancreatic cancer treatments.J Hematol Oncol. 2024 Oct 10;17(1):92. doi: 10.1186/s13045-024-01613-x. J Hematol Oncol. 2024. PMID: 39390609 Free PMC article. Review.
-
Role of chemokines in the crosstalk between tumor and tumor-associated macrophages.Clin Exp Med. 2023 Sep;23(5):1359-1373. doi: 10.1007/s10238-022-00888-z. Epub 2022 Sep 29. Clin Exp Med. 2023. PMID: 36173487 Free PMC article. Review.
-
Insulin/IGF Axis and the Receptor for Advanced Glycation End Products: Role in Meta-inflammation and Potential in Cancer Therapy.Endocr Rev. 2023 Jul 11;44(4):693-723. doi: 10.1210/endrev/bnad005. Endocr Rev. 2023. PMID: 36869790 Free PMC article. Review.
References
-
- Blair, A.B., Kim V.M., Muth S.T., Saung M.T., Lokker N., Blouw B., Armstrong T.D., Jaffee E.M., Tsujikawa T., Coussens L.M., et al. . 2019a. Dissecting the stromal signaling and regulation of myeloid cells and memory effector T cells in pancreatic cancer. Clin. Cancer Res. 25:5351–5363. 10.1158/1078-0432.CCR1078-018-4192 - DOI - PMC - PubMed
-
- Five Prime Therapeutics, Inc. 2020. Five Prime Therapeutics Provides Update on Phase 2 Trial of Cabiralizumab Combined with Opdivo® in Pancreatic Cancer. News release, February 18, 2020. Available at: https://www.businesswire.com/news/home/20200218005144/en/Five-Prime-Ther....
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
