

Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains
- PMID: 35408951
- PMCID: PMC8999060
- DOI: 10.3390/ijms23073590
Comparison of the Cisterna Maturation-Progression Model with the Kiss-and-Run Model of Intra-Golgi Transport: Role of Cisternal Pores and Cargo Domains
Abstract
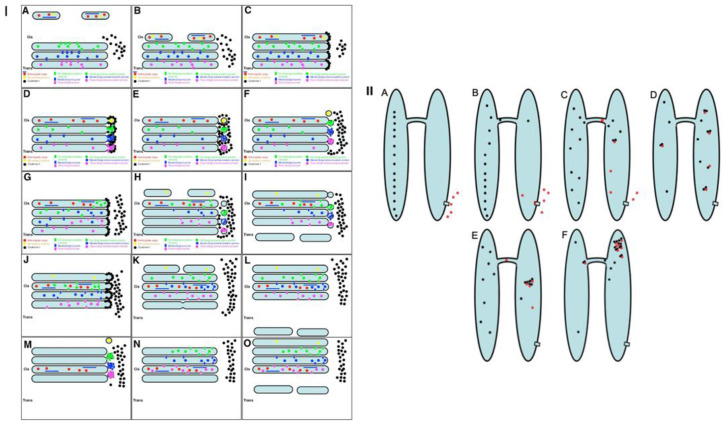
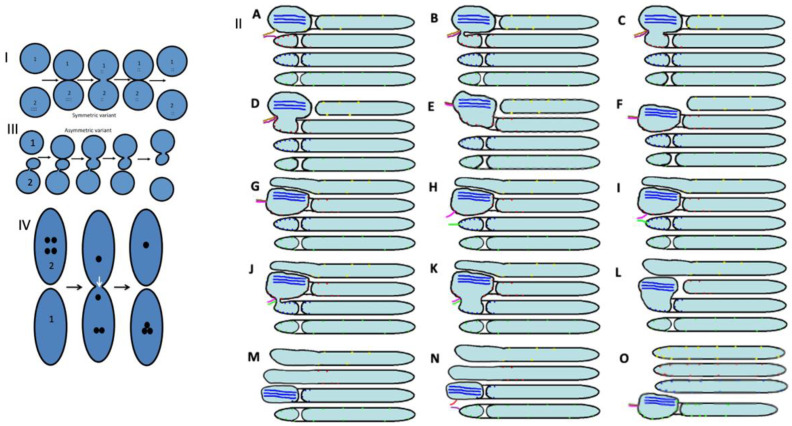
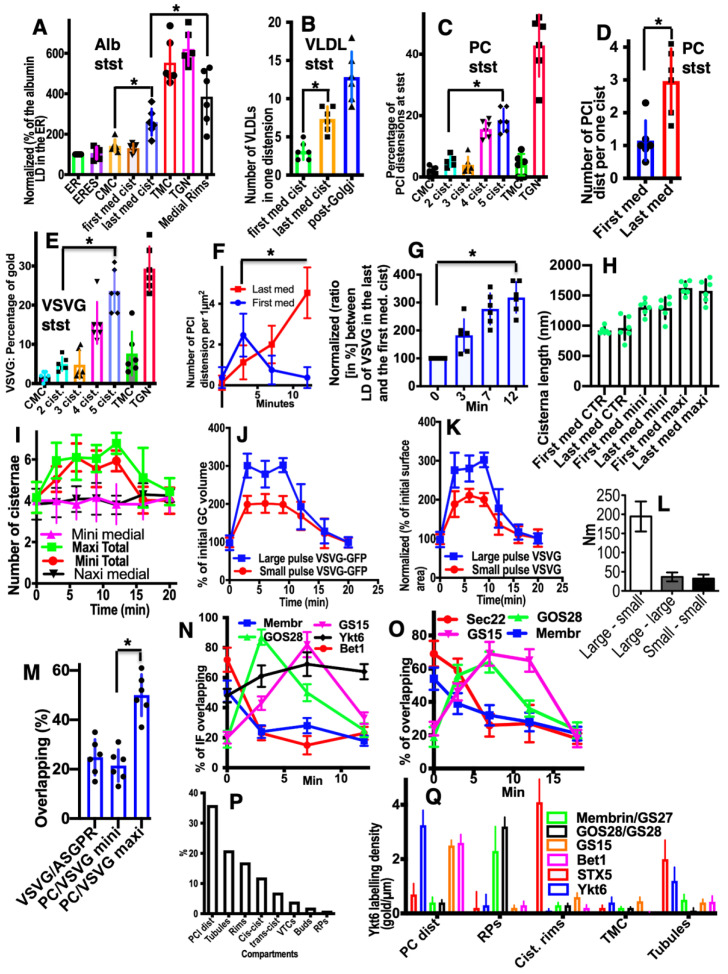
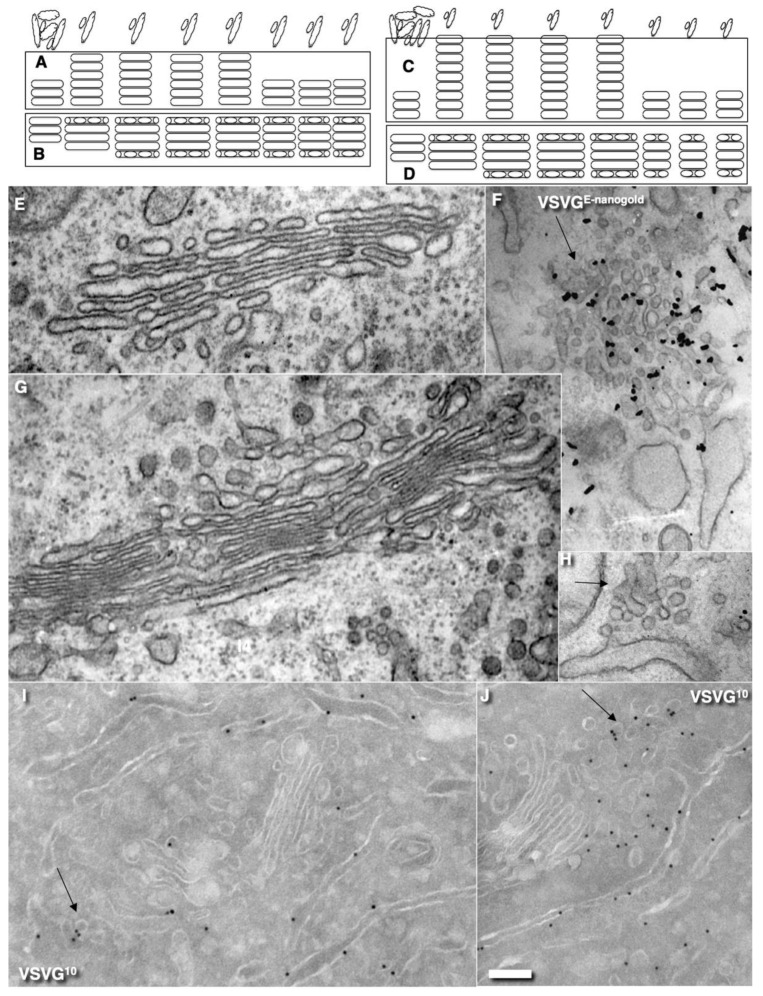
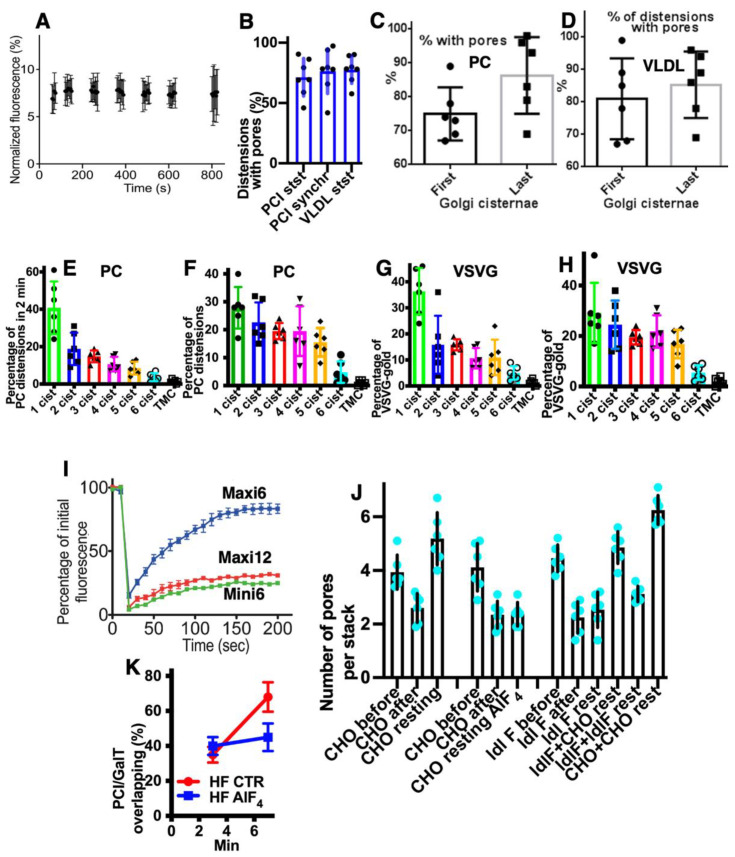
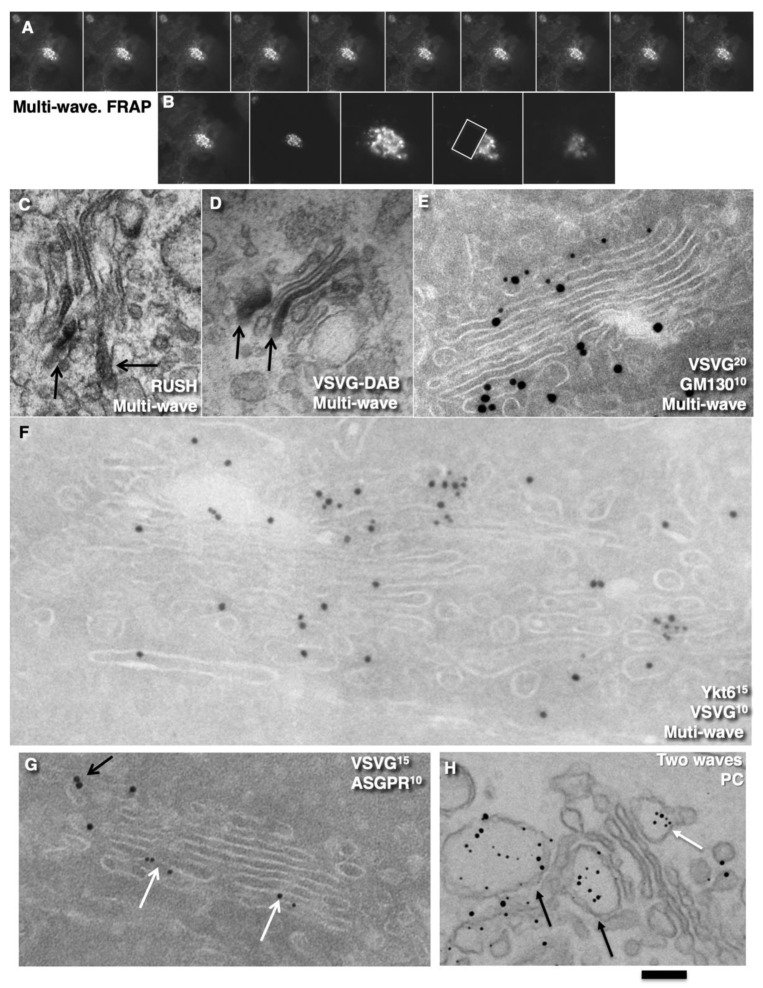
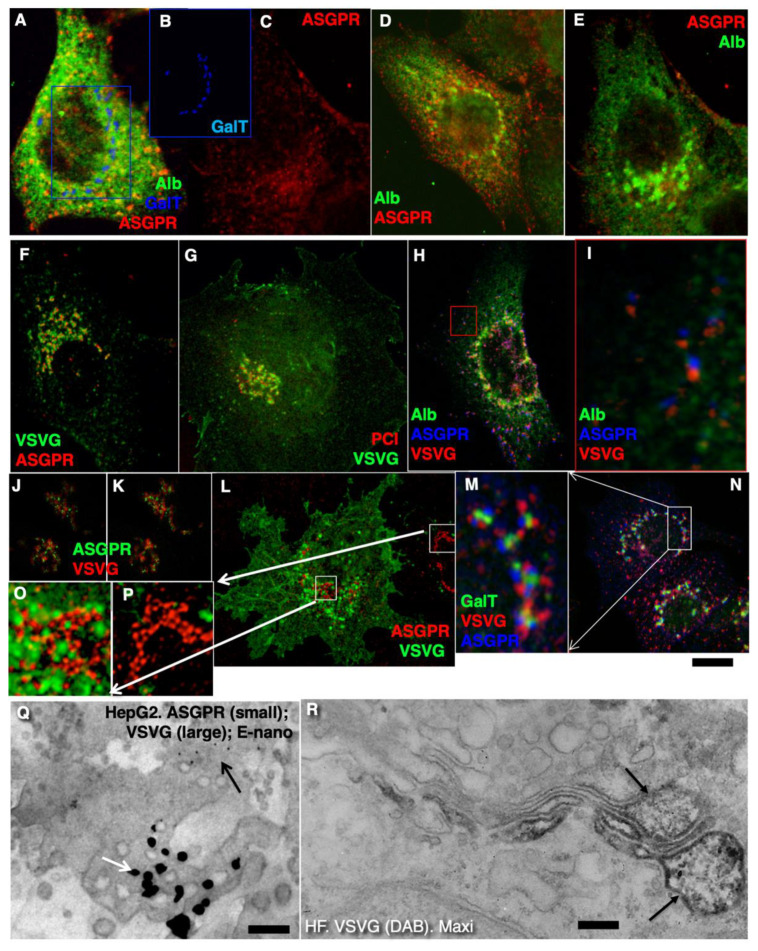
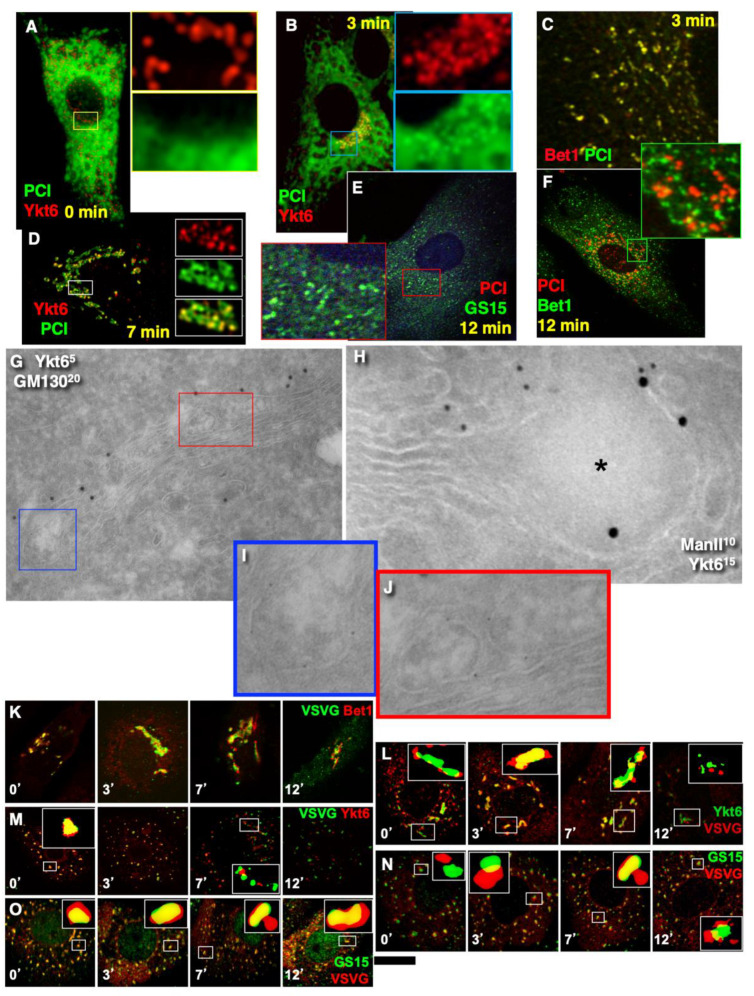
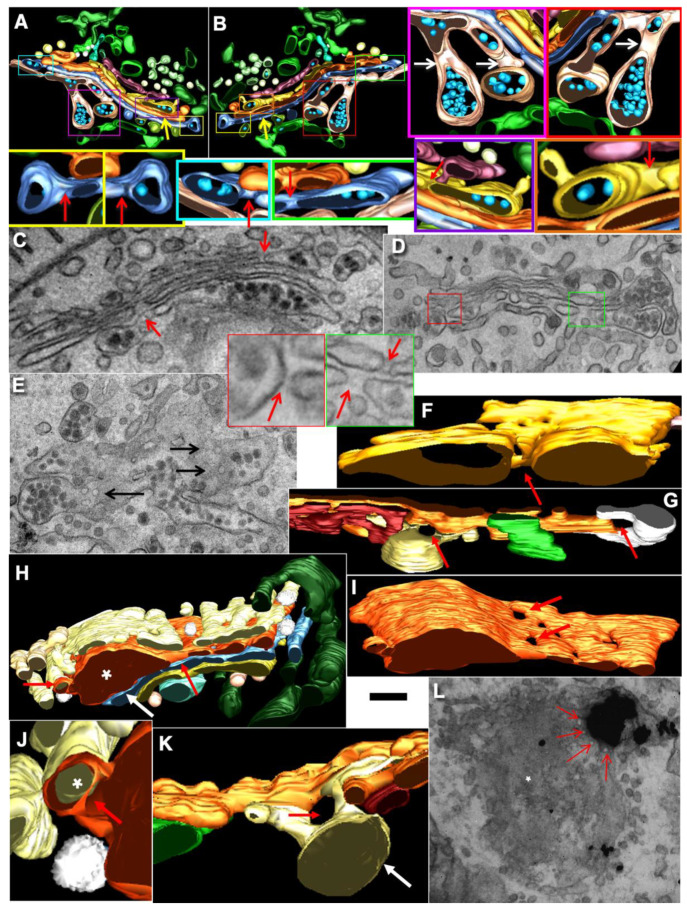
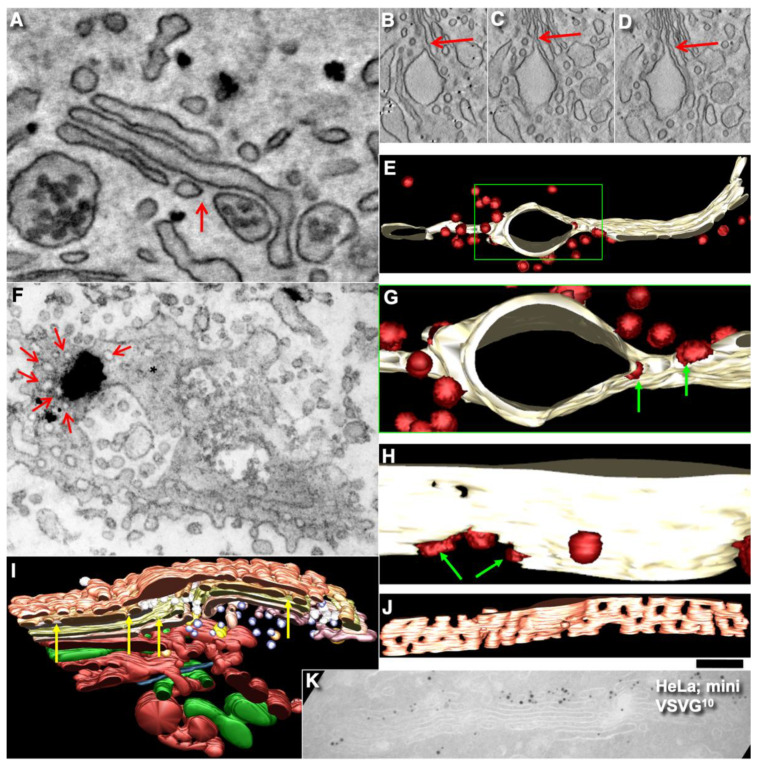
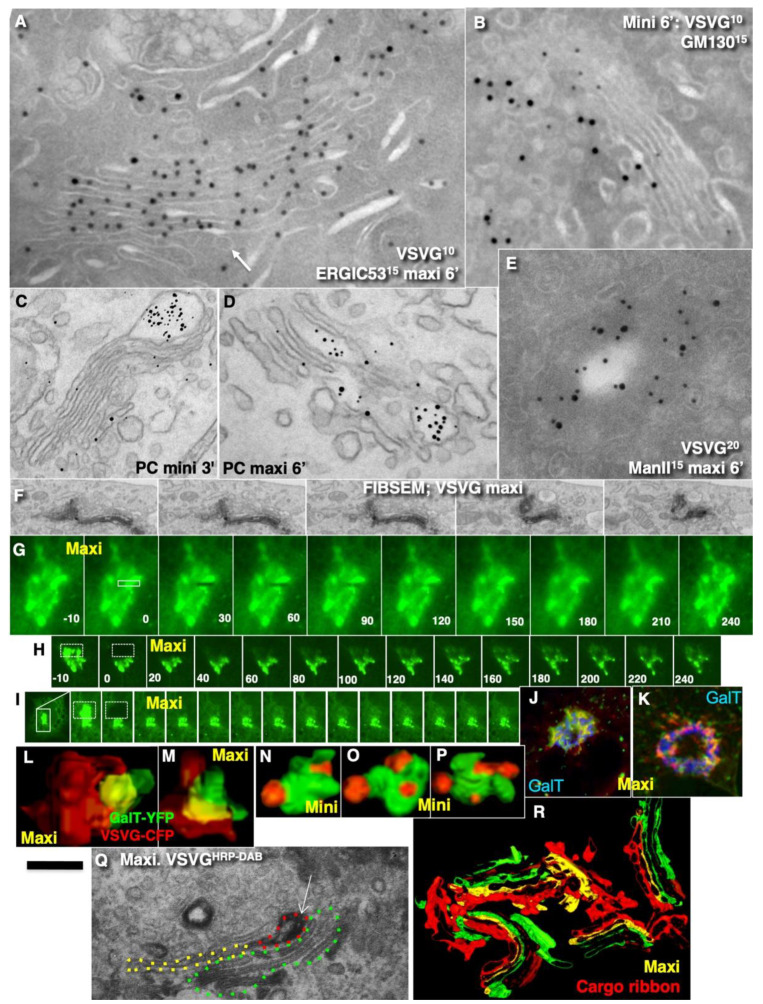
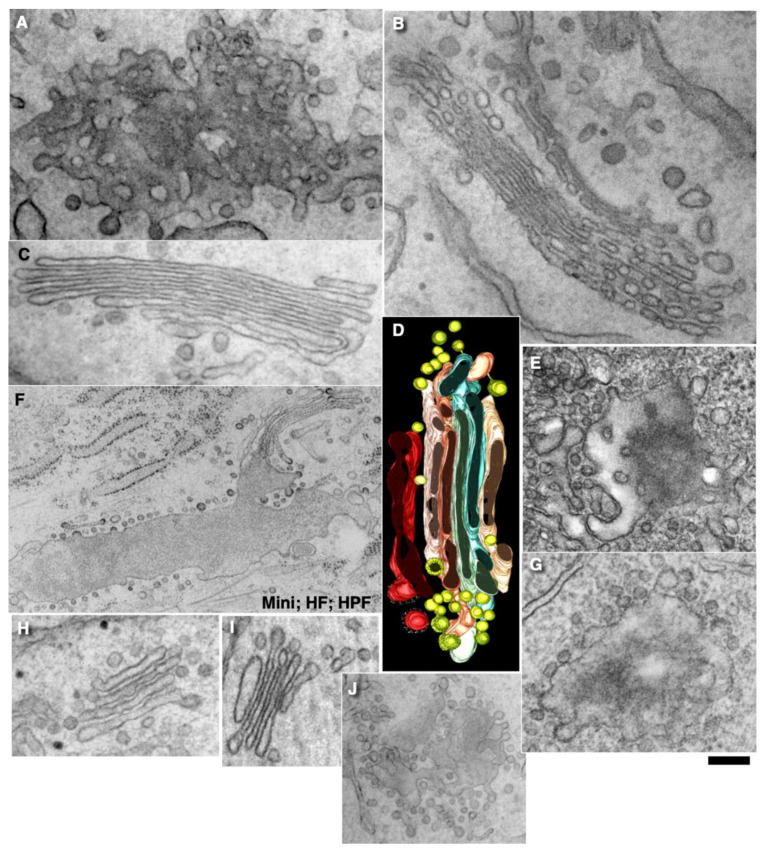
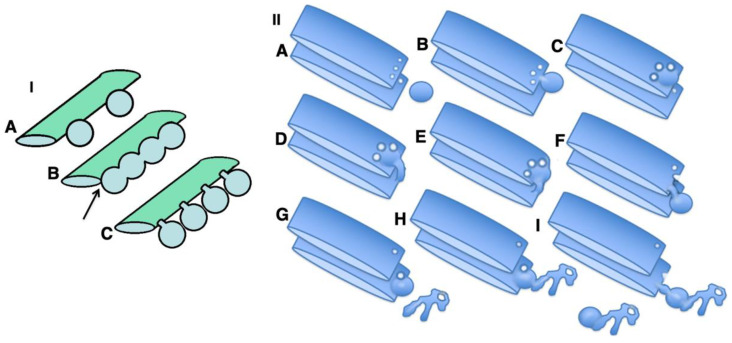
The Golgi complex is the central station of the secretory pathway. Knowledge about the mechanisms of intra-Golgi transport is inconsistent. Here, we compared the explanatory power of the cisterna maturation-progression model and the kiss-and-run model. During intra-Golgi transport, conventional cargoes undergo concentration and form cisternal distensions or distinct membrane domains that contain only one membrane cargo. These domains and distension are separated from the rest of the Golgi cisternae by rows of pores. After the arrival of any membrane cargo or a large cargo aggregate at the Golgi complex, the cis-Golgi SNAREs become enriched within the membrane of cargo-containing domains and then replaced by the trans-Golgi SNAREs. During the passage of these domains, the number of cisternal pores decreases. Restoration of the cisternal pores is COPI-dependent. Our observations are more in line with the kiss-and-run model.
Keywords: COPI; Golgi complex; Golgi dynamics; SNAREs; intracellular traffic; membrane fusion.
Conflict of interest statement
The authors declare that they have no conflict of interest. This article does not contain any study with humans, which was performed by the authors.
Figures


References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
