

Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma
- PMID: 35409129
- PMCID: PMC8998324
- DOI: 10.3390/ijms23073756
Identification, Isolation, and Characterization of Melanocyte Precursor Cells in the Human Limbal Stroma
Abstract
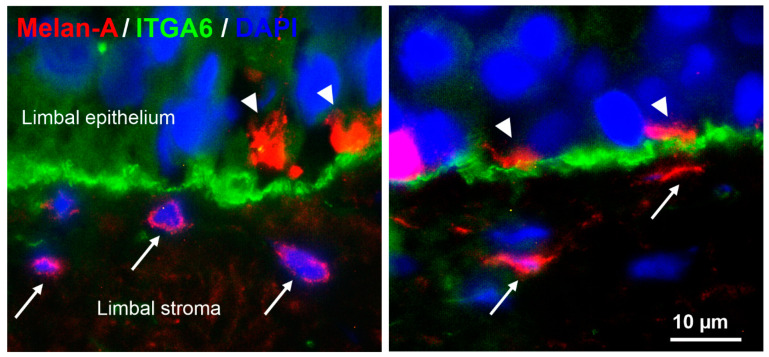
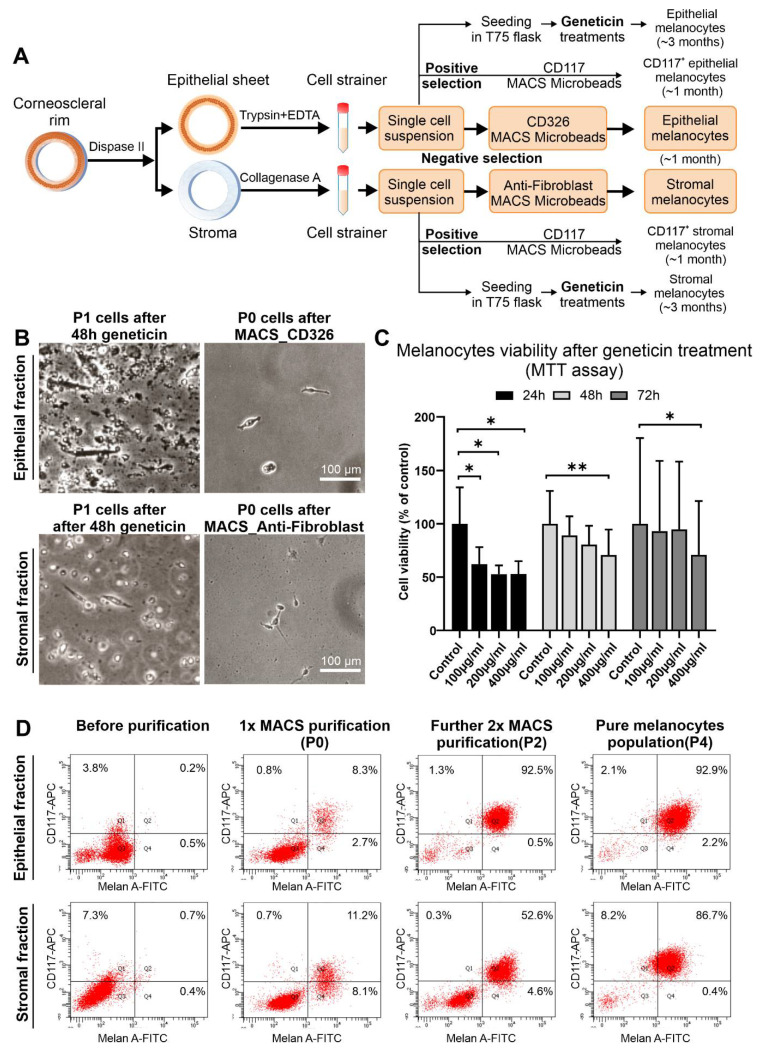
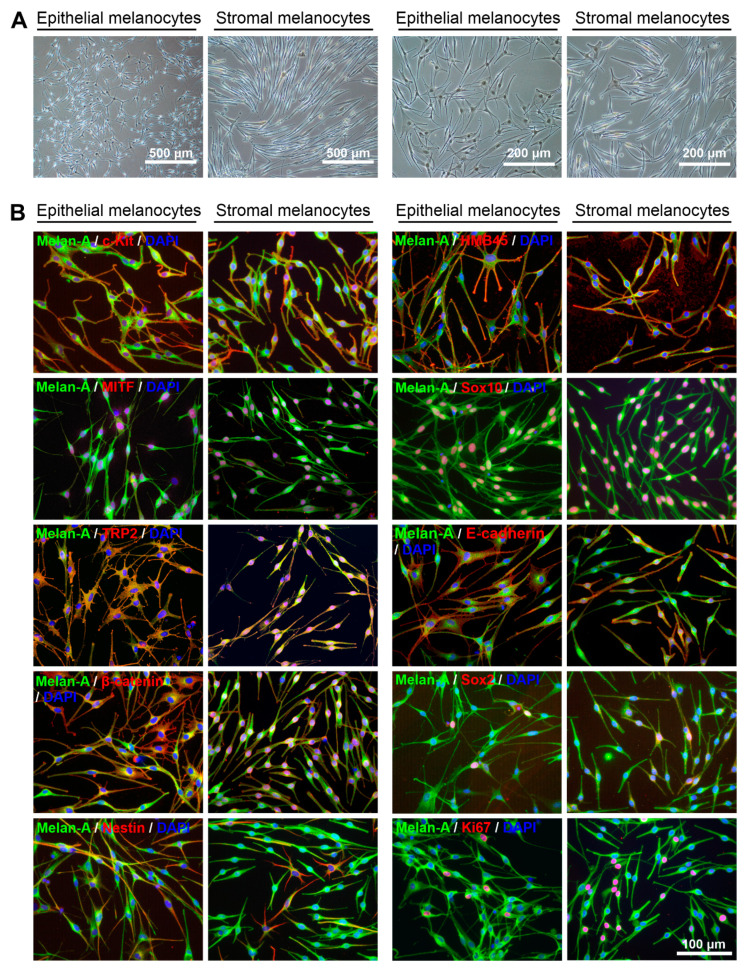
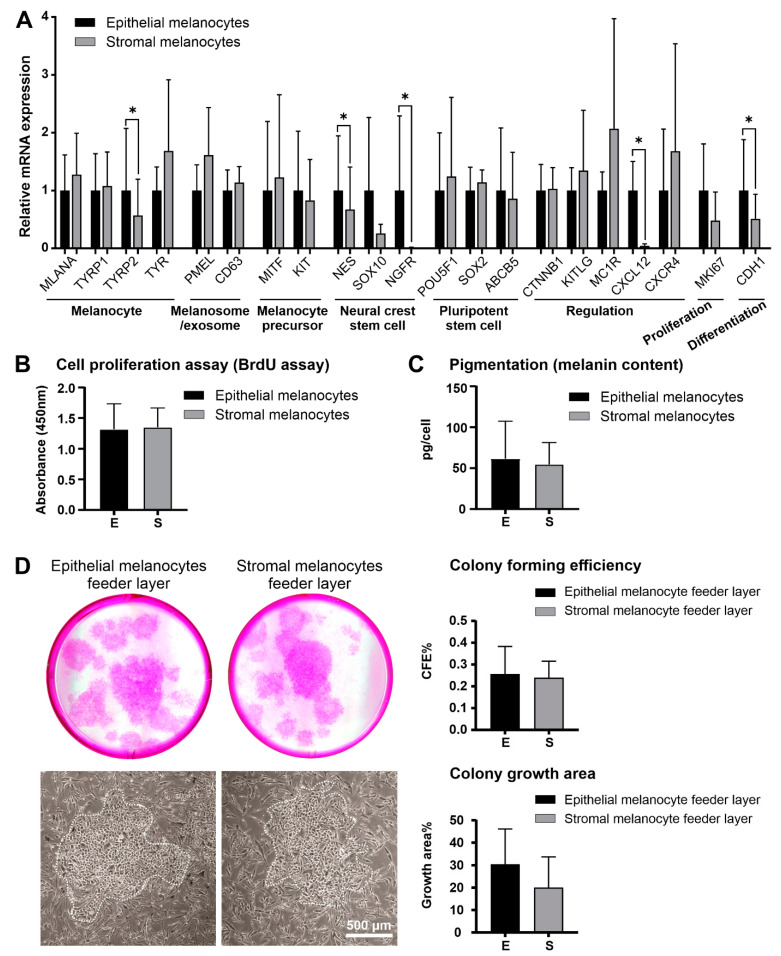
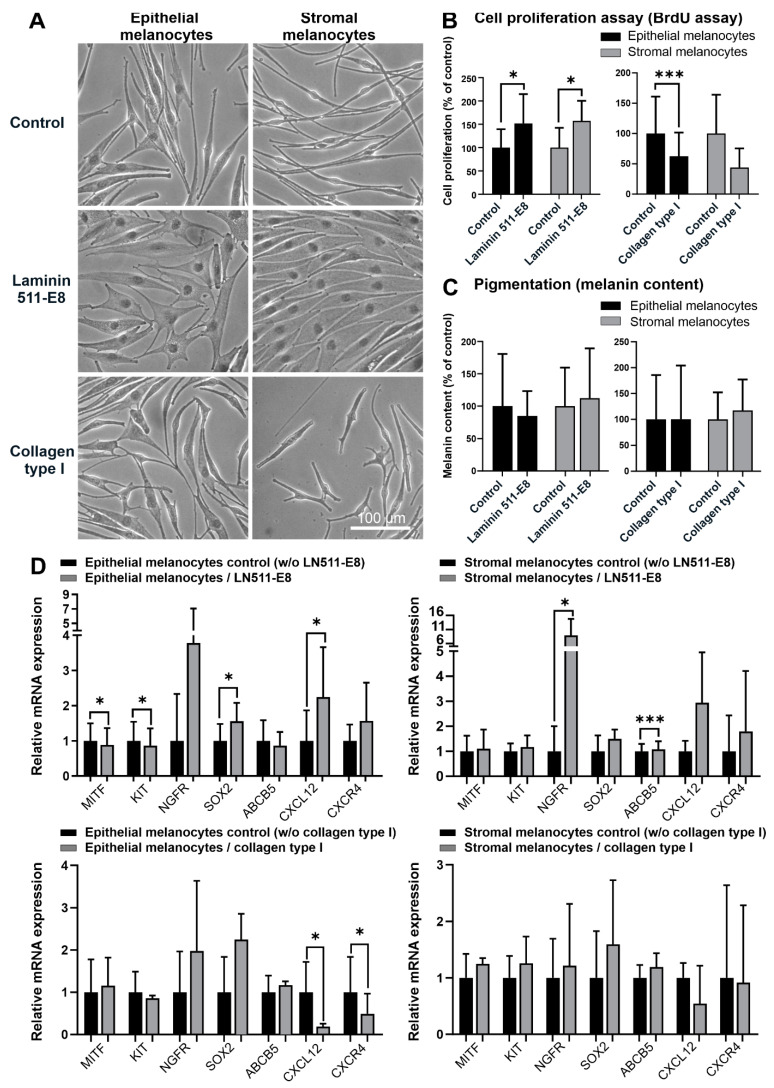
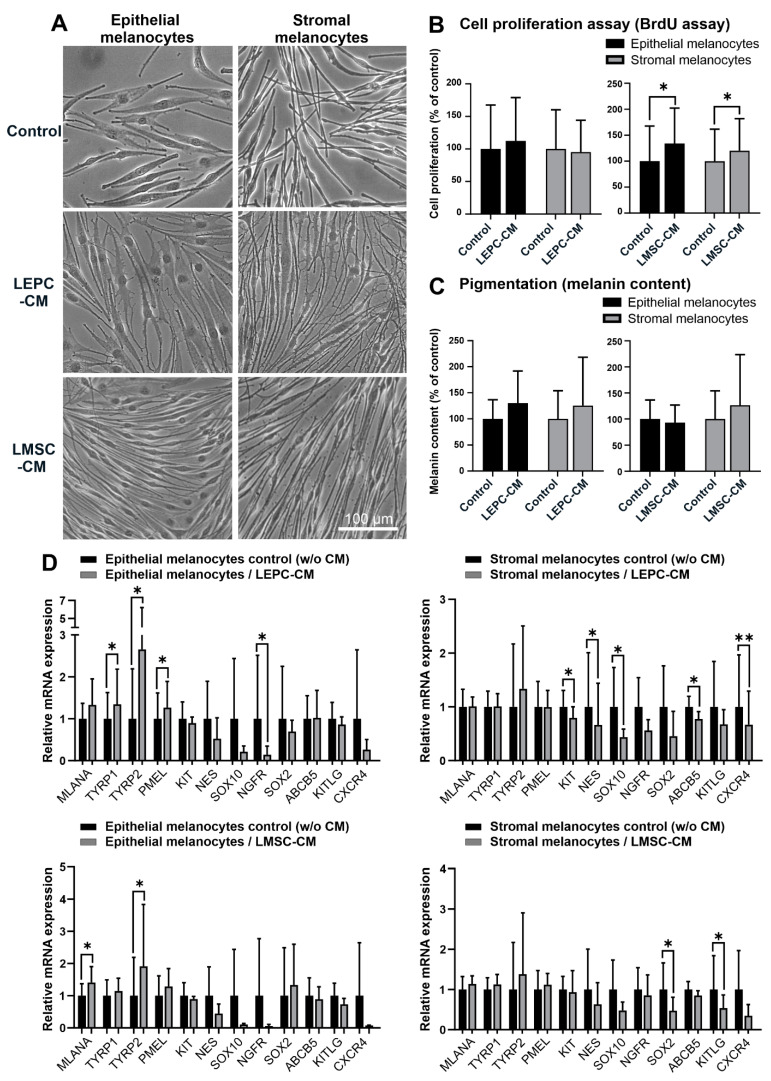
Given their vital role in the homeostasis of the limbal stem cell niche, limbal melanocytes have emerged as promising candidates for tissue engineering applications. This study aimed to isolate and characterize a population of melanocyte precursors in the limbal stroma, compared with melanocytes originating from the limbal epithelium, using magnetic-activated cell sorting (MACS) with positive (CD117/c-Kit microbeads) or negative (CD326/EpCAM or anti-fibroblast microbeads) selection approaches. Both approaches enabled fast and easy isolation and cultivation of pure limbal epithelial and stromal melanocyte populations, which differed in phenotype and gene expression, but exhibited similar functional properties regarding proliferative potential, pigmentation, and support of clonal growth of limbal epithelial stem/progenitor cells (LEPCs). In both melanocyte populations, limbus-specific matrix (laminin 511-E8) and soluble factors (LEPC-derived conditioned medium) stimulated melanocyte adhesion, dendrite formation, melanogenesis, and expression of genes involved in UV protection and immune regulation. The findings provided not only a novel protocol for the enrichment of pure melanocyte populations from limbal tissue applying easy-to-use MACS technology, but also identified a population of stromal melanocyte precursors, which may serve as a reservoir for the replacement of damaged epithelial melanocytes and an alternative resource for tissue engineering applications.
Keywords: cultivation; laminin 511-E8; limbal stem cell niche; limbal stem cells; limbal stroma; magnetic-activated cell sorting; melanocytes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Shortt A.J., Secker G.A., Munro P.M., Khaw P.T., Tuft S.J., Daniels J.T. Characterization of the Limbal Epithelial Stem Cell Niche: Novel Imaging Techniques Permit In Vivo Observation and Targeted Biopsy of Limbal Epithelial Stem Cells. Stem Cells. 2007;25:1402–1409. doi: 10.1634/stemcells.2006-0580. - DOI - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
