

Brain Metabolic Alterations in Alzheimer's Disease
- PMID: 35409145
- PMCID: PMC8998942
- DOI: 10.3390/ijms23073785
Brain Metabolic Alterations in Alzheimer's Disease
Abstract
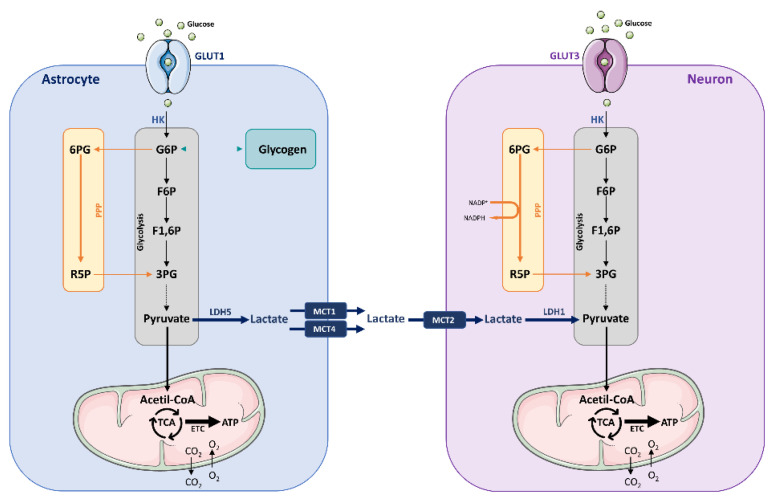
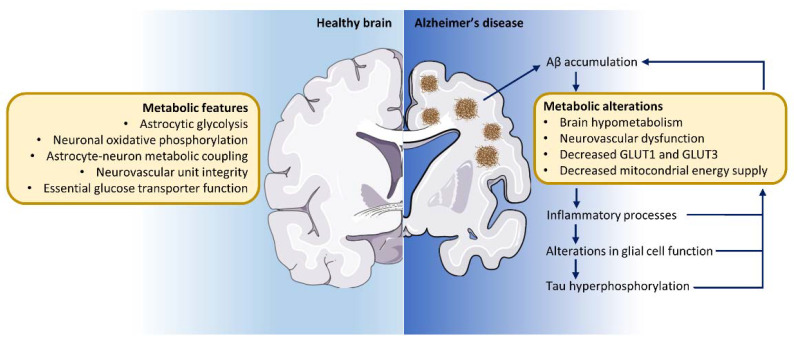
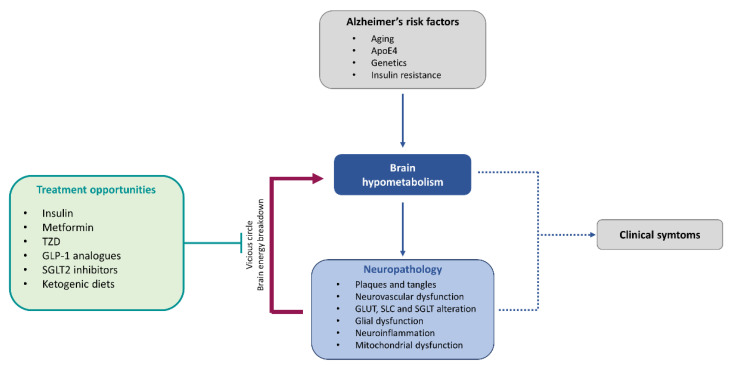
The brain is one of the most energy-consuming organs in the body. Satisfying such energy demand requires compartmentalized, cell-specific metabolic processes, known to be complementary and intimately coupled. Thus, the brain relies on thoroughly orchestrated energy-obtaining agents, processes and molecular features, such as the neurovascular unit, the astrocyte-neuron metabolic coupling, and the cellular distribution of energy substrate transporters. Importantly, early features of the aging process are determined by the progressive perturbation of certain processes responsible for adequate brain energy supply, resulting in brain hypometabolism. These age-related brain energy alterations are further worsened during the prodromal stages of neurodegenerative diseases, namely Alzheimer's disease (AD), preceding the onset of clinical symptoms, and are anatomically and functionally associated with the loss of cognitive abilities. Here, we focus on concrete neuroenergetic features such as the brain's fueling by glucose and lactate, the transporters and vascular system guaranteeing its supply, and the metabolic interactions between astrocytes and neurons, and on its neurodegenerative-related disruption. We sought to review the principles underlying the metabolic dimension of healthy and AD brains, and suggest that the integration of these concepts in the preventive, diagnostic and treatment strategies for AD is key to improving the precision of these interventions.
Keywords: GLUTs; astrocyte; astrocyte–neuron lactate shuttle (ANLS); glucose; hypometabolism; lactate; neurodegeneration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kety S.S. The general metabolism of the brain in vivo. In: Richter D., editor. Metabolism of the Nervous System. Peramon; London, UK: 1957. pp. 221–237.
-
- Sokoloff L., Reivich M., Kennedy C., Des Rosiers M.H., Patlak C.S., Pettigrew K.D., Sakurada O., Shinohara M. The [14C]deoxyglucose method for the measurement of local cerebral glucose utilization: Theory, procedure, and normal values in the conscious and anesthetized albino rat. J. Neurochem. 1977;28:897–916. doi: 10.1111/j.1471-4159.1977.tb10649.x. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
