

Inferring mammalian tissue-specific regulatory conservation by predicting tissue-specific differences in open chromatin
- PMID: 35410163
- PMCID: PMC8996547
- DOI: 10.1186/s12864-022-08450-7
Inferring mammalian tissue-specific regulatory conservation by predicting tissue-specific differences in open chromatin
Abstract
Background: Evolutionary conservation is an invaluable tool for inferring functional significance in the genome, including regions that are crucial across many species and those that have undergone convergent evolution. Computational methods to test for sequence conservation are dominated by algorithms that examine the ability of one or more nucleotides to align across large evolutionary distances. While these nucleotide alignment-based approaches have proven powerful for protein-coding genes and some non-coding elements, they fail to capture conservation of many enhancers, distal regulatory elements that control spatial and temporal patterns of gene expression. The function of enhancers is governed by a complex, often tissue- and cell type-specific code that links combinations of transcription factor binding sites and other regulation-related sequence patterns to regulatory activity. Thus, function of orthologous enhancer regions can be conserved across large evolutionary distances, even when nucleotide turnover is high.
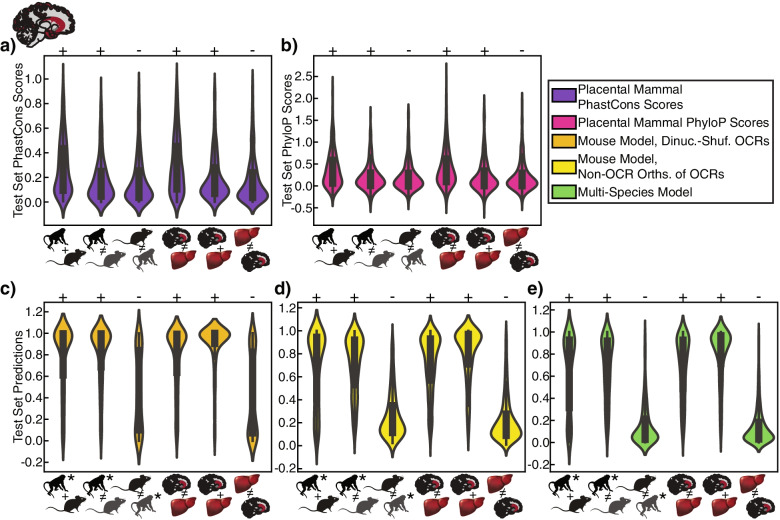
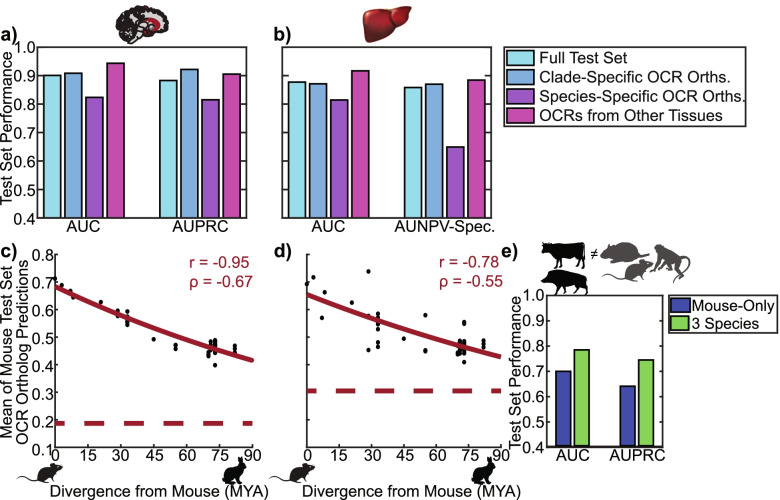
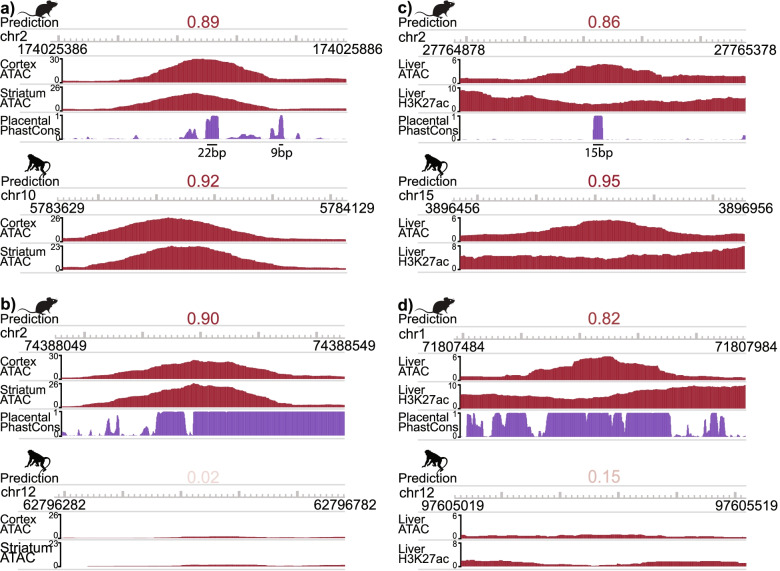
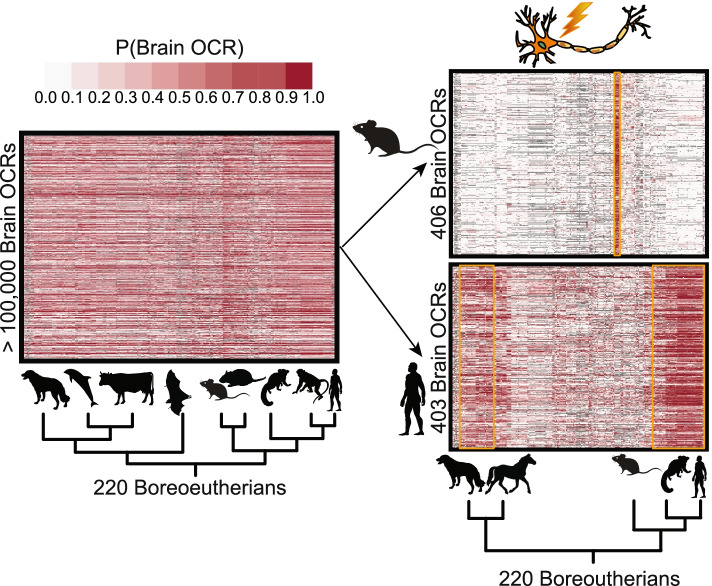
Results: We present a new machine learning-based approach for evaluating enhancer conservation that leverages the combinatorial sequence code of enhancer activity rather than relying on the alignment of individual nucleotides. We first train a convolutional neural network model that can predict tissue-specific open chromatin, a proxy for enhancer activity, across mammals. Next, we apply that model to distinguish instances where the genome sequence would predict conserved function versus a loss of regulatory activity in that tissue. We present criteria for systematically evaluating model performance for this task and use them to demonstrate that our models accurately predict tissue-specific conservation and divergence in open chromatin between primate and rodent species, vastly out-performing leading nucleotide alignment-based approaches. We then apply our models to predict open chromatin at orthologs of brain and liver open chromatin regions across hundreds of mammals and find that brain enhancers associated with neuron activity have a stronger tendency than the general population to have predicted lineage-specific open chromatin.
Conclusion: The framework presented here provides a mechanism to annotate tissue-specific regulatory function across hundreds of genomes and to study enhancer evolution using predicted regulatory differences rather than nucleotide-level conservation measurements.
Keywords: Enhancers; Gene expression evolution; Machine learning; Open chromatin prediction.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Bulik-Sullivan BK, Loh PR, Finucane HK, Ripke S, Yang J, Schizophrenia Working Group of the Psychiatric Genomics Consortium, Patterson N, Daly MJ, Price AL, Neale BM. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat Genet. 2015;47(3):291–5. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
