

Expansion of the prime editing modality with Cas9 from Francisella novicida
- PMID: 35410288
- PMCID: PMC8996390
- DOI: 10.1186/s13059-022-02644-8
Expansion of the prime editing modality with Cas9 from Francisella novicida
Abstract
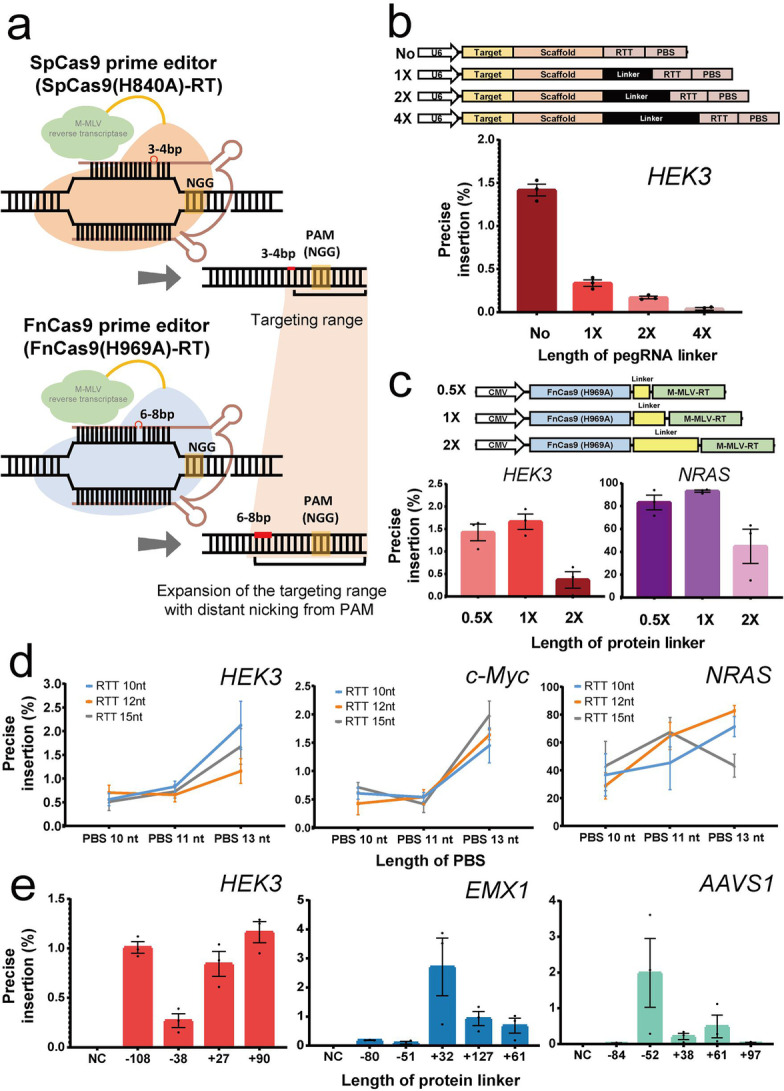
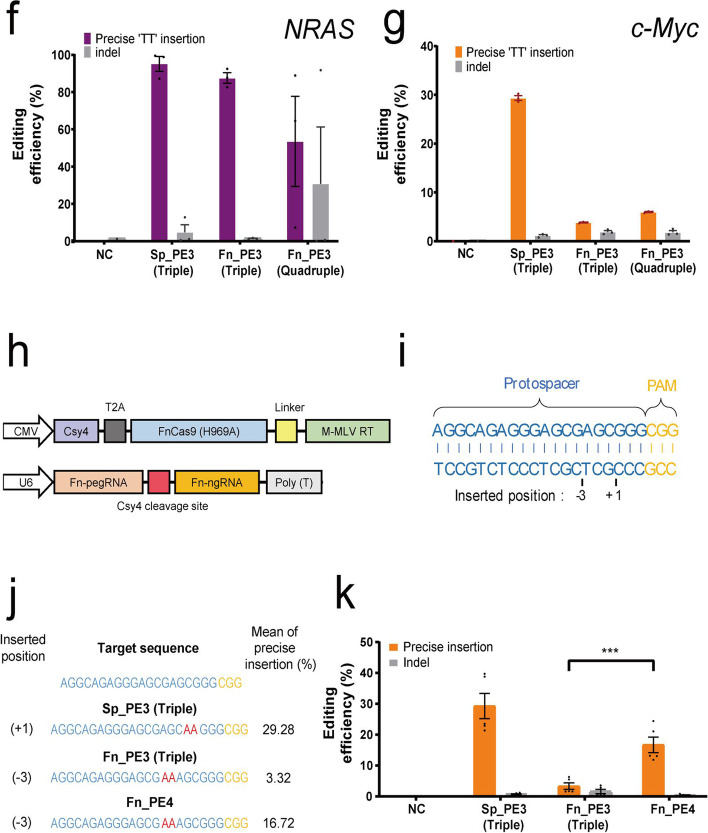
Prime editing can induce a desired base substitution, insertion, or deletion in a target gene using reverse transcriptase after nick formation by CRISPR nickase. In this study, we develop a technology that can be used to insert or replace external bases in the target DNA sequence by linking reverse transcriptase to the Francisella novicida Cas9, which is a CRISPR-Cas9 ortholog. Using FnCas9(H969A) nickase, the targeting limitation of existing Streptococcus pyogenes Cas9 nickase [SpCas9(H840A)]-based prime editing is dramatically extended, and accurate prime editing is induced specifically for the target genes in human cell lines.
Keywords: CRISPR-Cas9; Francisella novicida; Ortholog; Prime editing; Target expansion.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Chemello F, Chai AC, Li H, Rodriguez-Caycedo C, Sanchez-Ortiz E, Atmanli A, Mireault AA, Liu N, Bassel-Duby R, Olson EN. Precise correction of Duchenne muscular dystrophy exon deletion mutations by base and prime editing. Sci Adv. 2021;7(18):eabg4910. doi: 10.1126/sciadv.abg4910. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
