

MicroRNA networks in FLT3-ITD acute myeloid leukemia
- PMID: 35412895
- PMCID: PMC9169767
- DOI: 10.1073/pnas.2112482119
MicroRNA networks in FLT3-ITD acute myeloid leukemia
Abstract
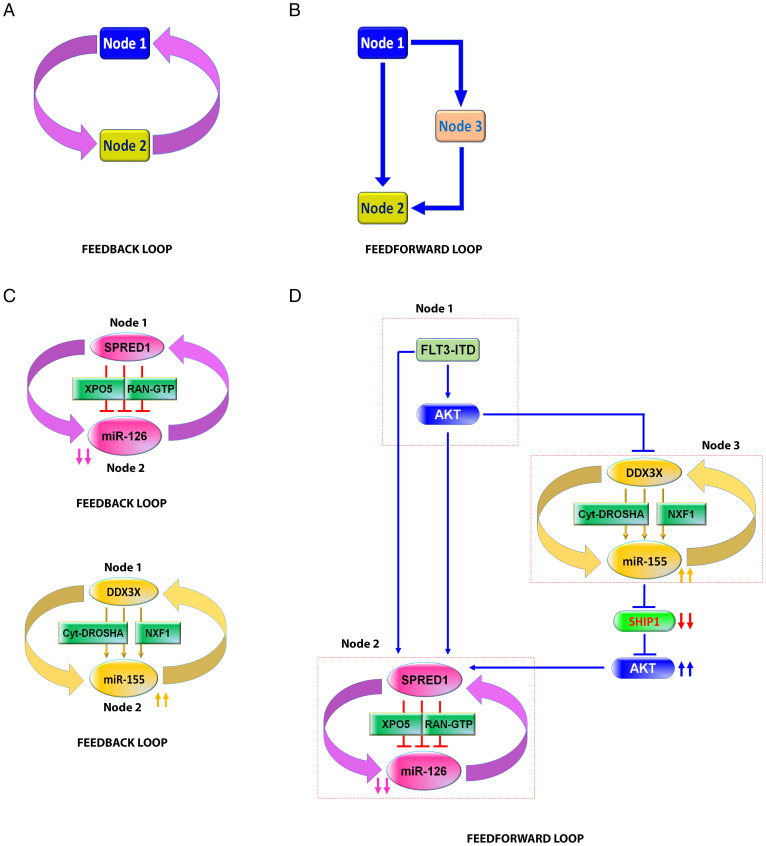
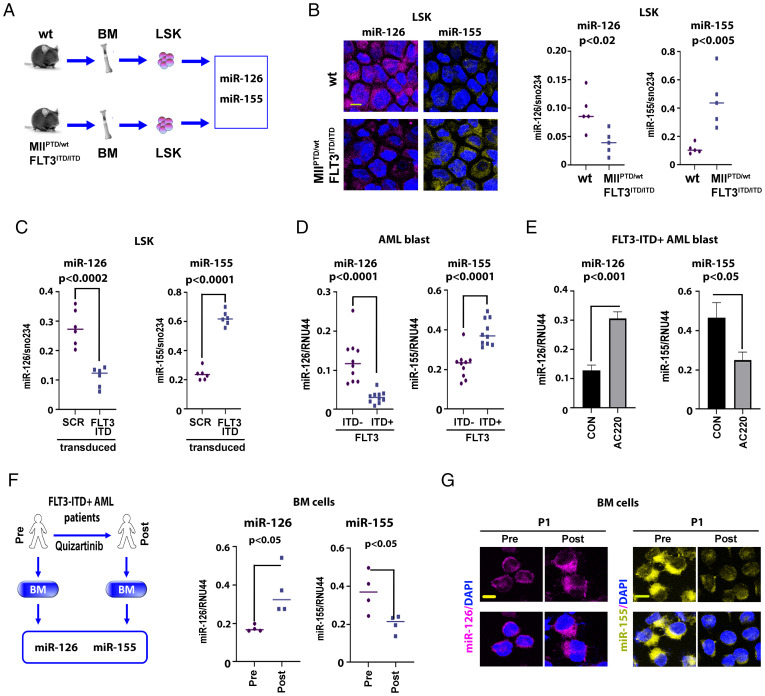
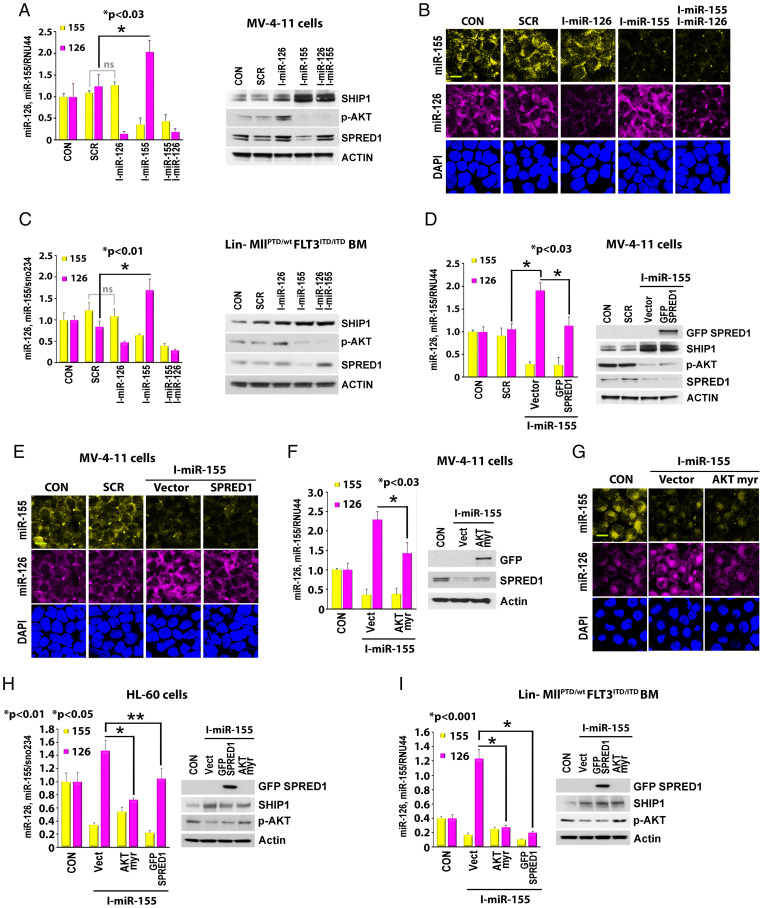
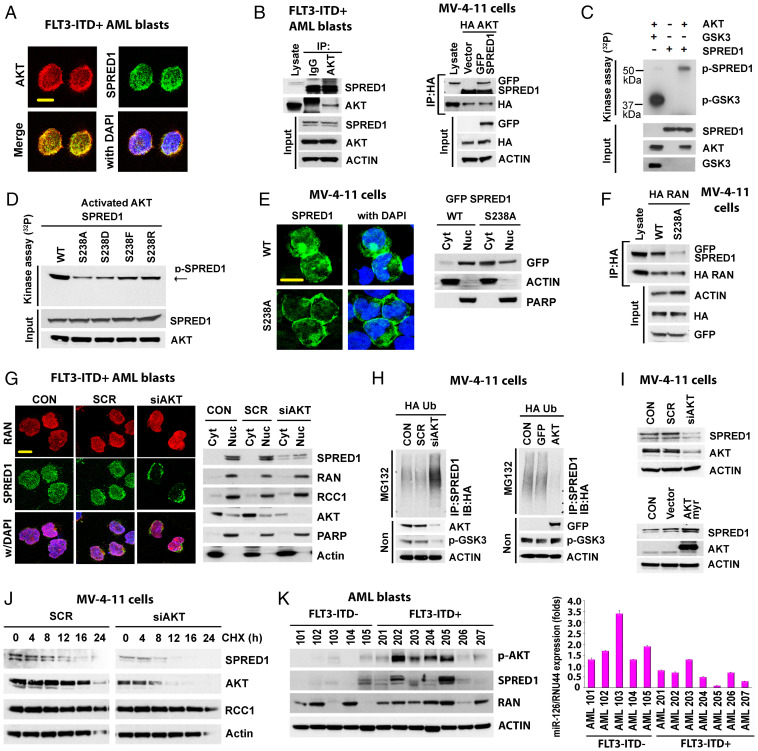
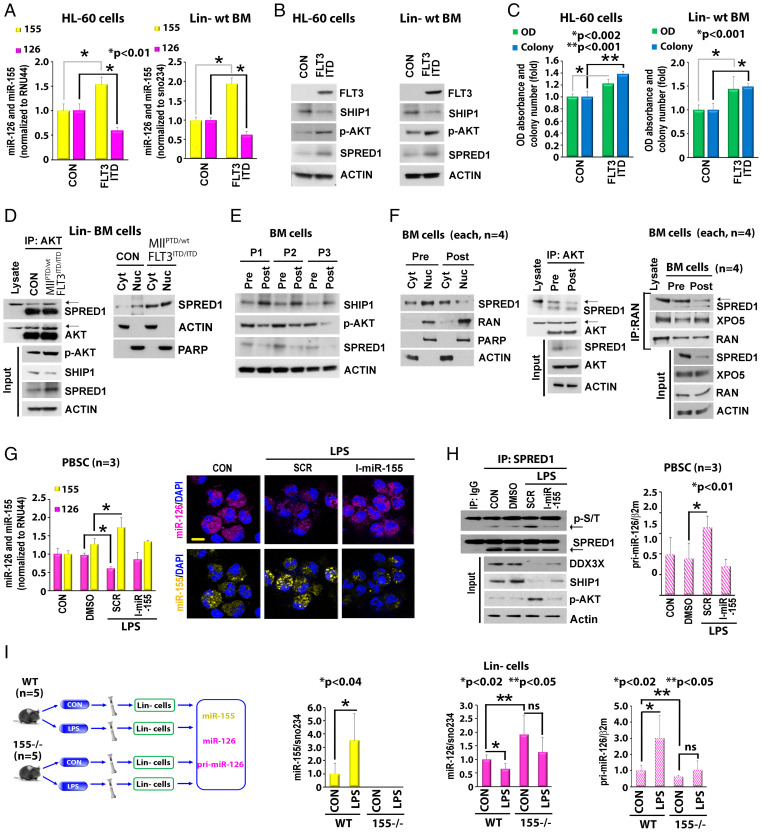
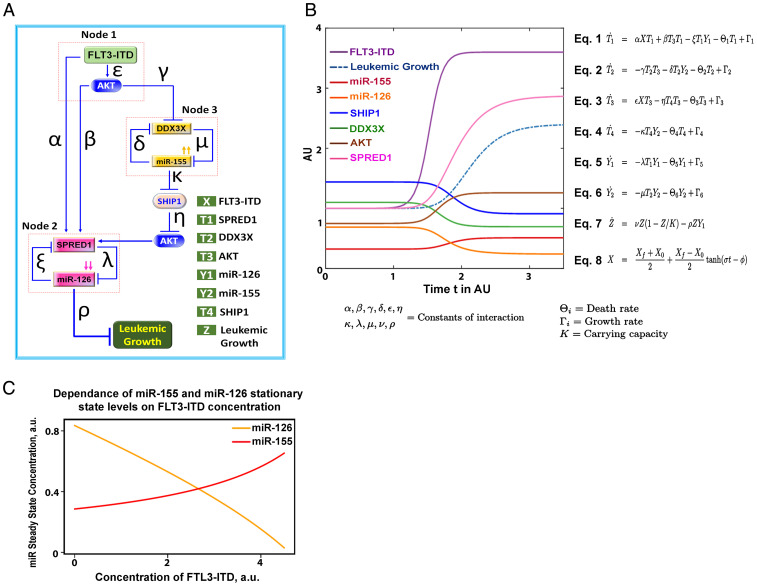
MiR-126 and miR-155 are key microRNAs (miRNAs) that regulate, respectively, hematopoietic cell quiescence and proliferation. Herein we showed that in acute myeloid leukemia (AML), the biogenesis of these two miRNAs is interconnected through a network of regulatory loops driven by the FMS-like tyrosine kinase 3-internal tandem duplication (FLT3-ITD). In fact, FLT3-ITD induces the expression of miR-155 through a noncanonical mechanism of miRNA biogenesis that implicates cytoplasmic Drosha ribonuclease III (DROSHA). In turn, miR-155 down-regulates SH2-containing inositol phosphatase 1 (SHIP1), thereby increasing phosphor-protein kinase B (AKT) that in turn serine-phosphorylates, stabilizes, and activates Sprouty related EVH1 domain containing 1 (SPRED1). Activated SPRED1 inhibits the RAN/XPO5 complex and blocks the nucleus-to-cytoplasm transport of pre-miR-126, which cannot then complete the last steps of biogenesis. The net result is aberrantly low levels of mature miR-126 that allow quiescent leukemia blasts to be recruited into the cell cycle and proliferate. Thus, miR-126 down-regulation in proliferating AML blasts is downstream of FLT3-ITD–dependent miR-155 expression that initiates a complex circuit of concatenated regulatory feedback (i.e., miR-126/SPRED1, miR-155/human dead-box protein 3 [DDX3X]) and feed-forward (i.e., miR-155/SHIP1/AKT/miR-126) regulatory loops that eventually converge into an output signal for leukemic growth.
Keywords: AKT; FLT3-ITD; acute myeloid leukemia; miR-126; miR-155.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
