

Strand asymmetry influences mismatch resolution during a single-strand annealing
- PMID: 35414014
- PMCID: PMC9001825
- DOI: 10.1186/s13059-022-02665-3
Strand asymmetry influences mismatch resolution during a single-strand annealing
Abstract
Background: Biases of DNA repair can shape the nucleotide landscape of genomes at evolutionary timescales. The molecular mechanisms of those biases are still poorly understood because it is difficult to isolate the contributions of DNA repair from those of DNA damage.
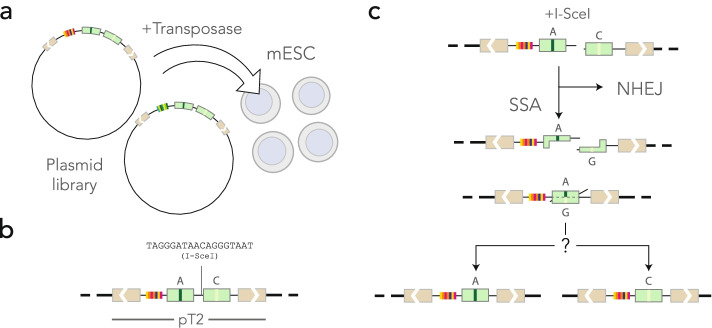
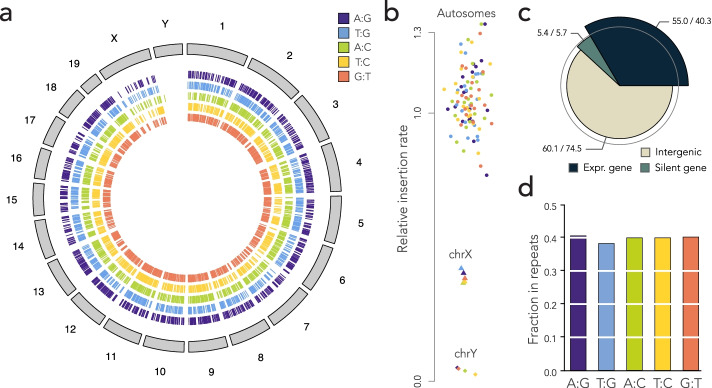
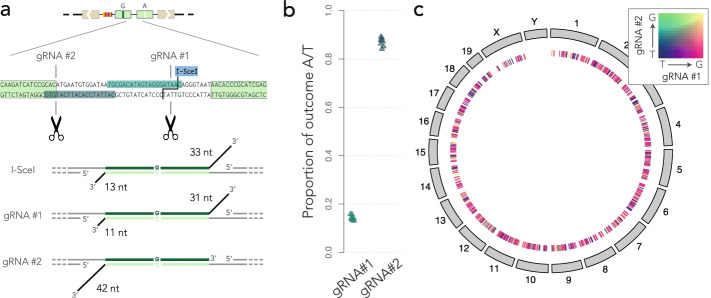
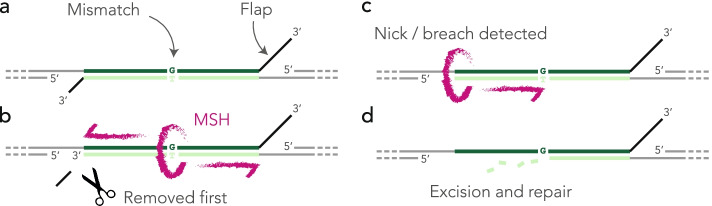
Results: Here, we develop a genome-wide assay whereby the same DNA lesion is repaired in different genomic contexts. We insert thousands of barcoded transposons carrying a reporter of DNA mismatch repair in the genome of mouse embryonic stem cells. Upon inducing a double-strand break between tandem repeats, a mismatch is generated if the break is repaired through single-strand annealing. The resolution of the mismatch showed a 60-80% bias in favor of the strand with the longest 3' flap. The location of the lesion in the genome and the type of mismatch had little influence on the bias. Instead, we observe a complete reversal of the bias when the longest 3' flap is moved to the opposite strand by changing the position of the double-strand break in the reporter.
Conclusions: These results suggest that the processing of the double-strand break has a major influence on the repair of mismatches during a single-strand annealing.
Keywords: Chromatin; Genome-wide technologies; Mismatch repair; Mouse embryonic stem cells; Single-strand annealing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
-
- Filipski J. Evolution of DNA sequence contributions of mutational bias and selection to the origin of chromosomal compartments. In: Obe G, editor. Advances in Mutagenesis Research. Berlin: Springer Berlin Heidelberg; 1990. pp. 1–54.
-
- Freese E. On the evolution of the base composition of DNA. J Theor Biol. 1962;3(1):82–101. doi: 10.1016/S0022-5193(62)80005-8. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
