

Genetic variation of putative myokine signaling is dominated by biological sex and sex hormones
- PMID: 35416774
- PMCID: PMC9094747
- DOI: 10.7554/eLife.76887
Genetic variation of putative myokine signaling is dominated by biological sex and sex hormones
Abstract
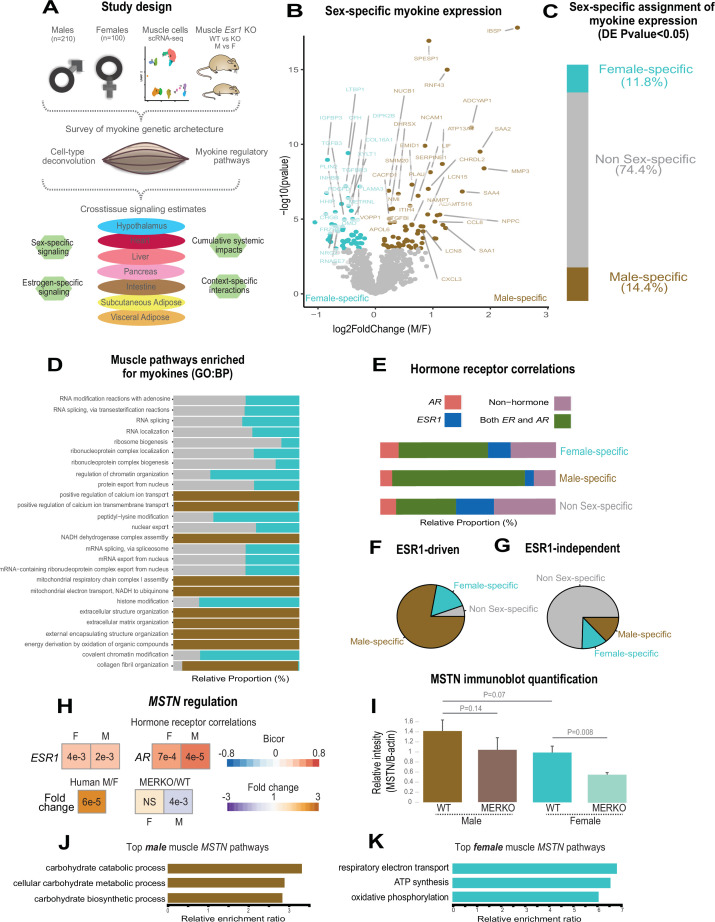
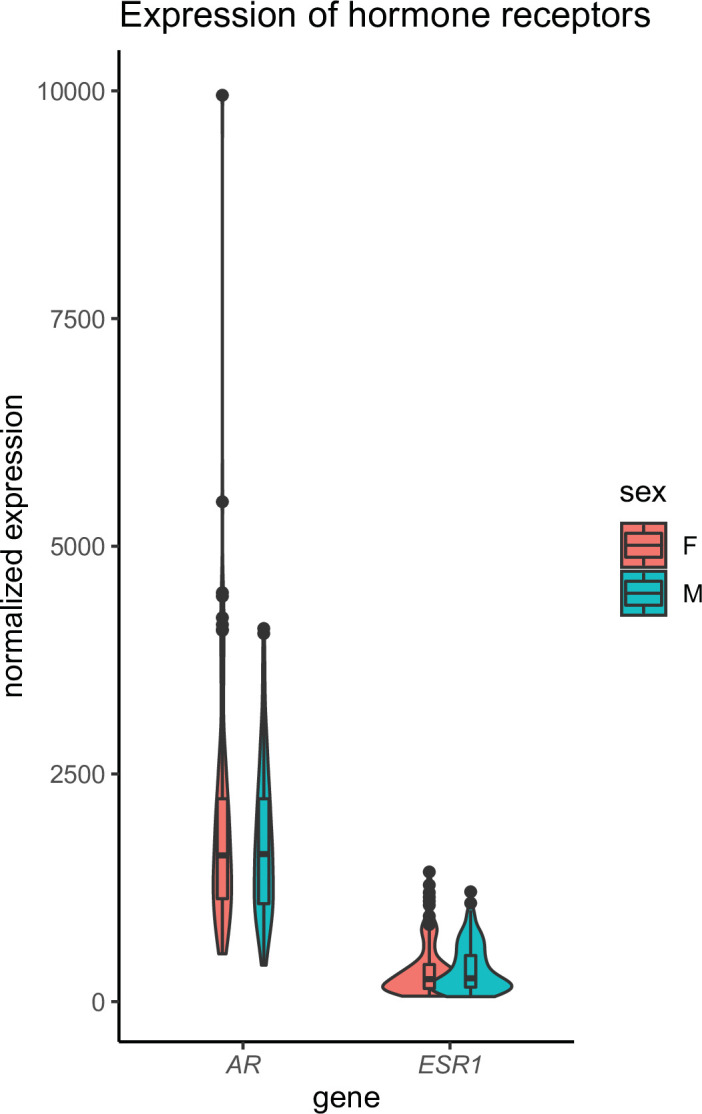
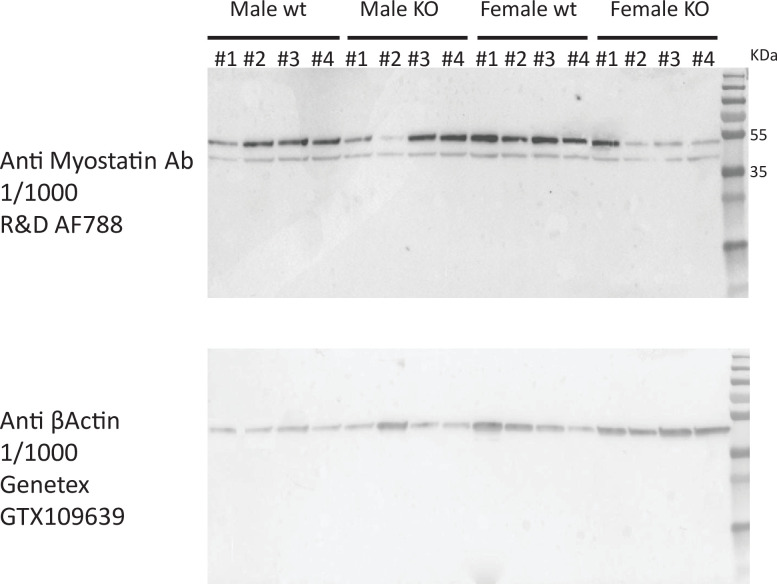
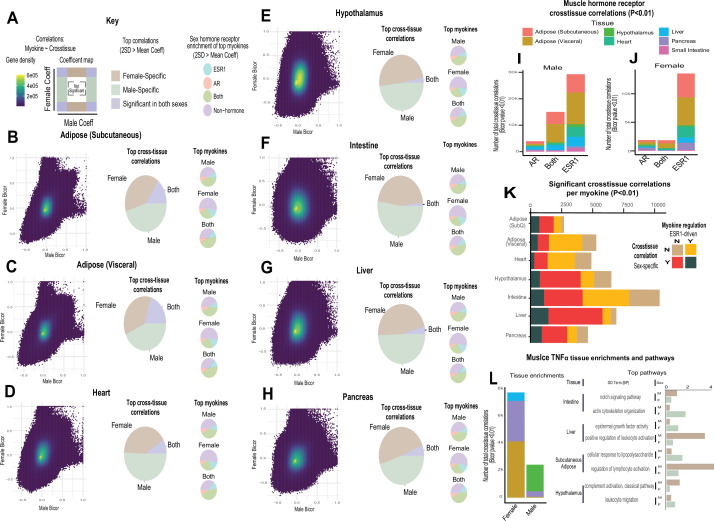
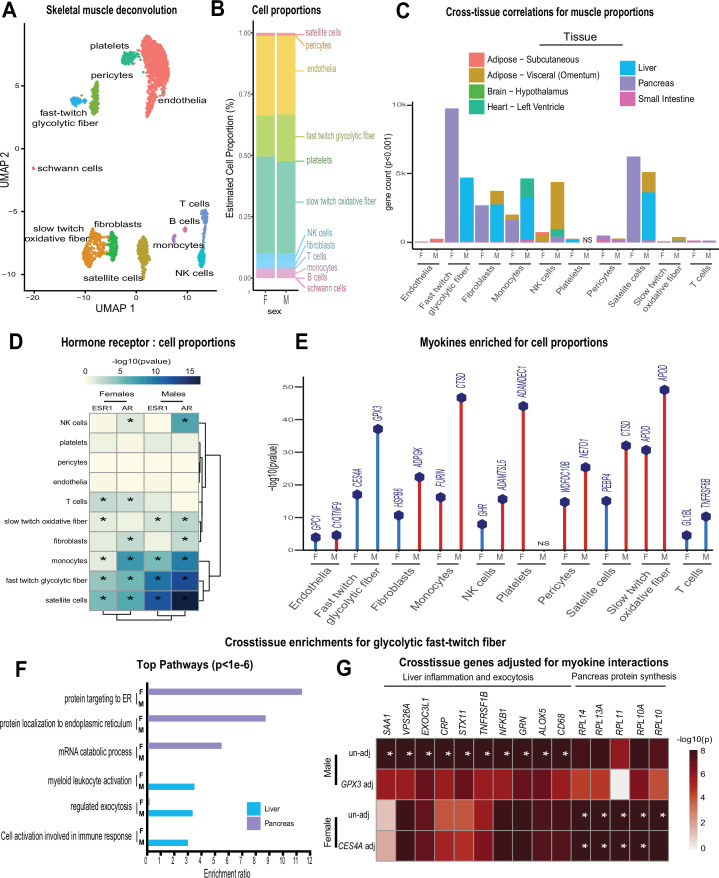
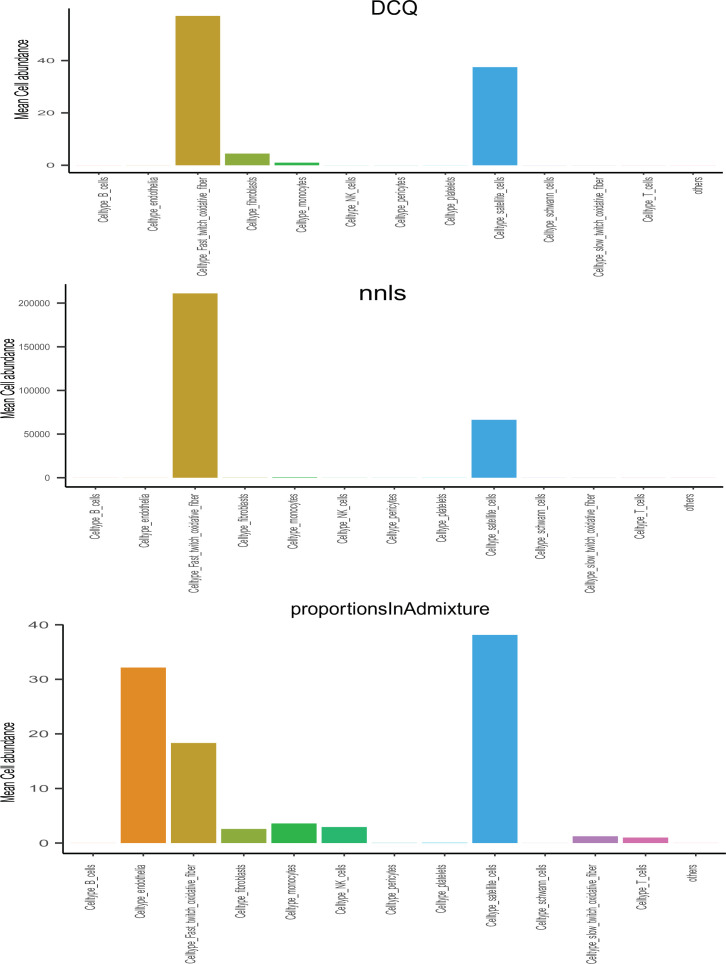
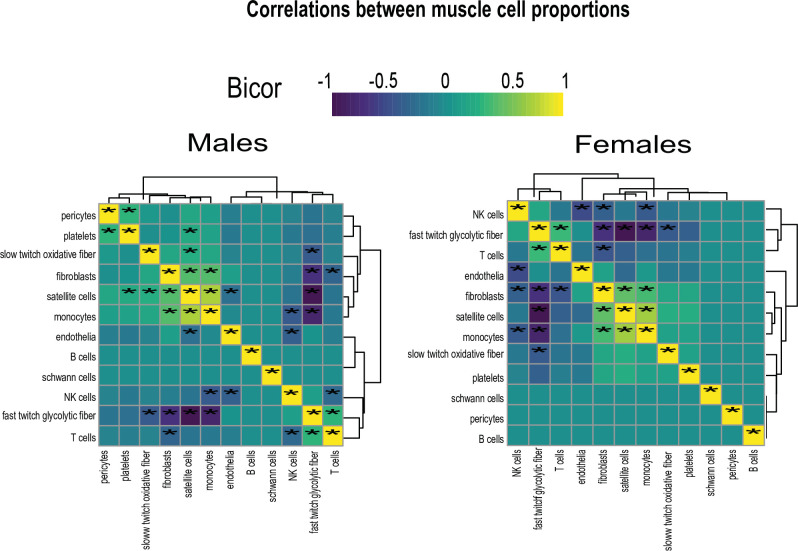
Skeletal muscle plays an integral role in coordinating physiological homeostasis, where signaling to other tissues via myokines allows for coordination of complex processes. Here, we aimed to leverage natural genetic correlation structure of gene expression both within and across tissues to understand how muscle interacts with metabolic tissues. Specifically, we performed a survey of genetic correlations focused on myokine gene regulation, muscle cell composition, cross-tissue signaling, and interactions with genetic sex in humans. While expression levels of a majority of myokines and cell proportions within skeletal muscle showed little relative differences between males and females, nearly all significant cross-tissue enrichments operated in a sex-specific or hormone-dependent fashion; in particular, with estradiol. These sex- and hormone-specific effects were consistent across key metabolic tissues: liver, pancreas, hypothalamus, intestine, heart, visceral, and subcutaneous adipose tissue. To characterize the role of estradiol receptor signaling on myokine expression, we generated male and female mice which lack estrogen receptor α specifically in skeletal muscle (MERKO) and integrated with human data. These analyses highlighted potential mechanisms of sex-dependent myokine signaling conserved between species, such as myostatin enriched for divergent substrate utilization pathways between sexes. Several other putative sex-dependent mechanisms of myokine signaling were uncovered, such as muscle-derived tumor necrosis factor alpha (TNFA) enriched for stronger inflammatory signaling in females compared to males and GPX3 as a male-specific link between glycolytic fiber abundance and hepatic inflammation. Collectively, we provide a population genetics framework for inferring muscle signaling to metabolic tissues in humans. We further highlight sex and estradiol receptor signaling as critical variables when assaying myokine functions and how changes in cell composition are predicted to impact other metabolic organs.
Keywords: biochemistry; chemical biology; computational biology; endocrinology; human; myokine; physiology; systems biology; systems genetics.
Plain language summary
The muscles that are responsible for voluntary movements such as exercise are called skeletal muscles. These muscles secrete proteins called myokines, which play roles in a variety of processes by interacting with other tissues. Essentially, myokines allow skeletal muscles to communicate with organs such as the kidneys, the liver or the brain, which is essential for the body to keep its metabolic balance. Some of the process myokines are involved include inflammation, cancer, the changes brought about by exercise, and even cognition. Despite the clear relevance of myokines to so many physiological outcomes, the way these proteins are regulated and their effects are not well understood. Genetic sex – specified by sex chromosomes in mammals – contributes to critical aspects of physiology. Specifically, many of the metabolic traits impacted by myokines show striking differences arising from hormonal or genetic interactions depending on the genetic sex of the subject being studied. It is therefore important to consider genetic sex when studying the effects of myokines on the body. Velez, Van et al. wanted to gain a better understanding of how skeletal muscles interact with metabolic tissues such as pancreas, liver and brain, taking genetic sex into consideration. To do this they surveyed human datasets for the correlations between the activity of genes that code for myokines, the composition of muscle cells, the signaling between muscles and metabolic tissues and genetic sex. Their results showed that, genetic sex and sex hormones predicted most of the effects of skeletal muscle on other tissues. For example, myokines from muscle were predicted to be more impactful on liver or pancreas, depending on whether individuals were male or female, respectively. The results of Velez, Van et al. illustrate the importance of considering the effects of genetic sex and sexual hormones when studying metabolism. In the future, these results will allow other researchers to design sex-specific experiments to be able to gather more accurate information about the mechanisms of myokine signaling.
© 2022, Velez et al.
Conflict of interest statement
LV, CV, TM, ZZ, CJ, AH, MS No competing interests declared
Figures
References
-
- Chella Krishnan K, Vergnes L, Acín-Pérez R, Stiles L, Shum M, Ma L, Mouisel E, Pan C, Moore TM, Péterfy M, Romanoski CE, Reue K, Björkegren JLM, Laakso M, Liesa M, Lusis AJ. Sex-specific genetic regulation of adipose mitochondria and metabolic syndrome by Ndufv2. Nature Metabolism. 2021;3:1552–1568. doi: 10.1038/s42255-021-00481-w. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- dbGaP/phs000424.v8.p2
- Actions
